Table 1. Change of D10 values under different irradiation conditions of E. coli S2 and injury ratio of cell membrane enumerated by different cultivation media.
 |
実験方法
1. 供試菌株
2. ラジカル捕捉剤
3. 照射
4. 生残菌数の測定
実験結果
1. E. coli S2 株の放射線感受性に関与するフリーラジカルの影響
2. 各菌株の放射線感受性に及ぼす培養基の影響
Escherichia coli O157:H7 や Salmonella enteritidis、Staphylococcus aureus 等の食品由来の病原性非芽胞細菌は比較的放射線感受性が高く、冷蔵食肉中では 1 〜 3kGy で殺菌可能である。しかし、これらの菌の放射線殺菌効果は生残菌数を測定する培養基の種類によって異なることがあり、殺菌線量評価の上で問題を生じる可能性がある。一方、加熱殺菌や凍結障害、等の場合にも培養基の種類によって細菌の生残率が著しく異なることが報告されている1)。加熱や凍結、等の微生物障害の場合には DNA 鎖の切断も重要な要因であるが、細胞膜の障害も同時に起こり、このことが培養基の種類による生残率の差に大きく関与している。
微生物の放射線障害は基本的に DNA 鎖の切断であり、放射線障害にはヒドロキシラジカル (OH) やスーパーオキシドラジカル (O2-) が関与していると報告されている2 〜 4)。しかし、放射線による E. coli の突然変異発生率は紫外線と大差がなく、生存率 10-6 において突然変異発生率は 0.02 〜 0.26% にすぎない5)。一方、微生物に対する放射線障害の機構及び培養基の影響の有無を明らかにすることも微生物学的安全性評価の上で重要と思われる。
本研究では OH ラジカル等のフリーラジカルと酸素の放射線感受性に与える影響及び培養基の影響を比較し、放射線による細胞傷害の機構について検討した。
供試菌株は下水汚泥から分離された Escherichia coli S2 株、放射線耐性の E. coli B/r 株、牛肉より分離され E. coli O157:H7 A4-2 株との共生株で同じ放射線感受性を示す A4-1 株6)、Salmonella enteritidis YK-2 株、Staphylococcus aureus H12 株を用いた。これらの菌株は Nutrient broth で 30℃・16 時間振とう培養し、遠心分離後に 0.067M 燐酸緩衝液 (pH7.0) で 2 回洗浄し、同緩衝液に菌数を約 1 x 109 cfu/ml となるように懸濁した。
放射線によって生成する細胞内外の OH ラジカルをほぼ完全に捕捉する目的で 2M グリセリン2) を菌懸濁液に添加した。また、細胞外の OH ラジカルを捕捉する目的で 1.5% ポリエチレングリコール−600 を添加した。蟻酸は O2- ラジカル生成の目的で添加し4)、40mM を燐酸緩衝液に加え pH を 7.0 に調整した。
菌懸濁液約 5ml をガス置換可能なパイレックスガラス試験管に入れ、窒素ガス (N2)、酸素ガス (O2)、一酸化二窒素ガス (N2O) を各 2 分間通気し、ガス飽和下で密封した。ガンマ線源は 10 万キュリー・コバルト−60 を用い、1.2kGy/h の位置で照射し、照射位置の線量率は Fricke 鉄線量計であらかじめ測定しておいた。
照射後の菌液の希釈は滅菌生理食塩水 (0.01% Tween 20 含有) で行い、生残菌数の測定には Tryptic soy agar を細胞膜傷害回復用培養基として用いた。細胞膜の障害検出用培養基としては、E. coli 及び S. enteritidis に MacConkey agar を用い、St. aureus には 7% NaCl Tryptic soy agar を用いた1)。生残菌数は平板培地表面に塗布し、30℃・1 晩培養して出現してくるコロニー数から測定した。細胞膜の傷害が生じた細胞はデソキシコレートや NaCl に感受性となると報告されており1)、MacConkey agar と 0.2% デソキシコレート含有 Tryptic soy agar が同じ感受性を示すことをあらかじめ確認しておいた。
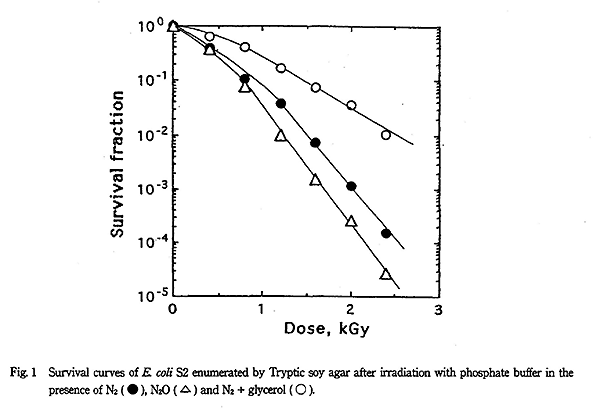
Tryptic soy agar で得られた E. coli S2 株の放射線感受性は Fig. 1 に示すようになり、異なった照射条件により D10 値に著しい差が認められた。すなわち、窒素ガス飽和下で得られた D10 値は 0.56kGy であったのに対し、窒素ガス+グリセリン共存下での D10 値は 0.96kGy になった。一方、窒素ガス+ポリエチレングリコール共存下での D10 値は窒素ガス飽和下と同じになった。また、一酸化二窒素ガス飽和下での D10 値は 0.46kGy であり、Fig. 2 のフリーラジカル生成機構で示すように OH ラジカルが約 2 倍に増加するにもかかわらず感受性促進効果はわずかしか認められなかった。
これに対し、酸素ガス飽和下での D10 値は 0.19kGy であり、窒素ガス飽和に比べ 3 倍近い感受性促進効果が認められた。
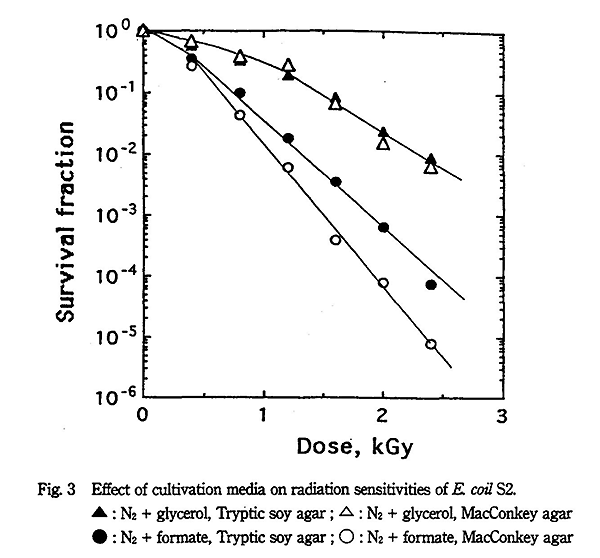
E. coli S2 株の放射線感受性を窒素ガス+グリセリンまたは蟻酸共存下で Tryptic soy agar 及び MacConkey agar で比較したところ、Fig. 3 に示すようにグリセリン共存下では培養基による差が全く認められなかったのに対し、蟻酸共存下では著しい差が認められた。MacConkey agar で感受性が増加するのは細胞膜障害によりデソキシコレート感受性となるためであり、蟻酸共存下で生成した O2- ラジカルが細胞膜障害に大きく関与していることを示している。一方、酸素ガス飽和下でも培養基による感受性に大きな差が認められた。すなわち、E. coli S2 株の Tryptic soy agar と MacConkey agar で得られた D10 値の差は Table 1 に示すように酸素ガス飽和、酸素ガス+グリセリン及び窒素ガス+蟻酸共存下で約 16% となるのに対し、窒素ガス飽和及び一酸化二窒素ガス飽和では約 7%、窒素ガス+グリセリン共存下で 0% となった。
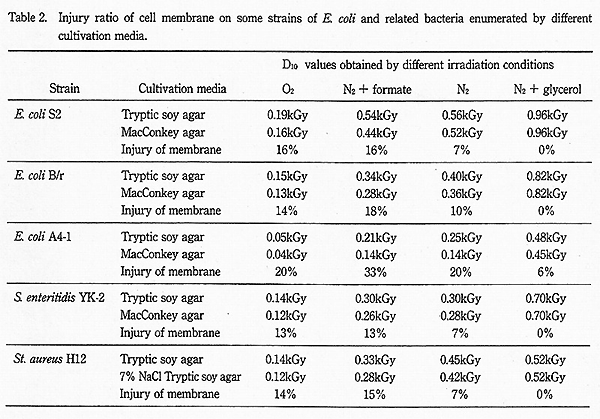
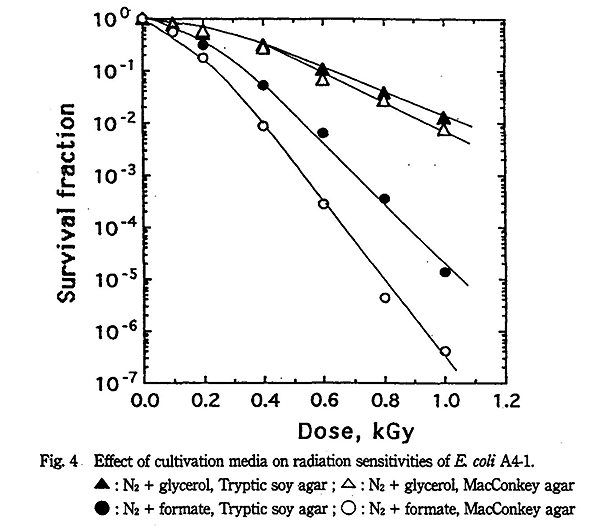
また、Table 2 に示すように、E. coli B/r、S. enteritidis YK-2 及び St. aureus H12 株の培養基による感受性変化の傾向も E. coli S2 株と似ており、窒素ガス+グリセリン共存下で培養基による差が認められなかったのに対し、酸素ガス飽和下または窒素ガス+蟻酸共存下では培養基による感受性に著しい差が認められた。一方、放射線感受性株の E. coli A4-1 株では Fig. 4 に示すように、窒素ガス+グリセリン共存下でも培養基による感受性に差が認められ、窒素ガス+蟻酸共存下での培養基による D10 値の差は 33% になった。また、酸素ガス飽和下と窒素ガス飽和下では培養基による D10 値の差は各々 20% となり、両者は同じ値を示した。
E. coli S2 株等の放射線感受性で明らかなように、グリセリンは細胞内外の OH ラジカルを捕捉するのに対し、ポリエチレングリコールは細胞外の OH ラジカルのみを捕捉することを示している。このことは、放射線で生成した OH ラジカルは細胞内で生成した場合にのみ放射線感受性に関与することを示している。Samni ら3) は T7 バクテリオファージの放射線感受性について検討し、ファージの不活性化には OH ラジカルが大きく関与し、O2- ラジカルは不活性化にほとんど関与しないと報告している。一方、Kim ら4) は S. typhimurium の放射線感受性には OH ラジカルばかりでなく O2- ラジカルが寄与し、O2- ラジカルは細胞内の損傷を促進すると報告している。従来、O2- ラジカルの生物組織等に対する反応は OH ラジカルに変換されてから作用すると考えられてきたが、Samni らの報告3) や Kim らの報告4) からは OH ラジカルへの変換はほとんどなく、O2- ラジカルは直接酸化反応に関与していることを示唆している。
本研究の結果では、E. coli 等の殺菌効果には DNA 鎖の切断ばかりでなく、細胞膜の障害も関与していることが明らかになった。しかも、放射線による細胞膜の障害は放射線感受性の高い株ほど起こりやすく、細胞膜の障害に最も寄与するのは O2- ラジカルであり、分子状酸素や OH ラジカルも関与することを示している。しかし本研究の結果では、OH ラジカルの生成量が増加しても放射線感受性の著しい増大は認められず、酸素ガス飽和下で感受性が著しく増大するのは分子状酸素が DNA 損傷に関与するためと思われる4)。一方、OH や O2- ラジカルが影響しにくい系では細胞膜の障害が全く認められない株が多かったが、放射線感受性株の E. coli A4-1 株では細胞膜の障害がわずかながら認められた。このことは、OH ラジカルがほとんど生成しにくい放射線の直接作用でも細胞膜の障害が起こる可能性を示しており、E. coli A4-1 株のように細胞膜障害の影響が現れやすい株と E. coli S2 株などのような細胞傷害が現れにくい株があることを示しているように思われる。また、この結果は微生物の放射線感受性は DNA の修復機構だけでは説明できないことを示唆している。しかし、本研究の結果からすると肉類や魚貝類中に存在する病原菌の多くは溶存酸素がほとんど存在しない環境下にあり、しかも天然のラジカル捕捉剤と共存しているため、細胞膜の障害はほとんど起こらず、培養基の種類による殺菌線量の評価に大きな影響はないものと思われる。
1) 森地敏樹 : 防菌防徹, 17, 541-550 (1989).
2) D. Ewing and K. E. Damasker : Biochem. Biophys. Res. Commun., 207, 515-520 (1995).
3) A. Samni, M. Chevion and G. Czapski : Radiat. Res., 99, 562-572 (1984).
4) A. Y. Kim and D. W. Thyer : Radiat. Res., 144, 36-42 (1995).
5) 瀧上真智子, 伊藤 均 : 食品照射, 30, 11-16 (1995).
6) 伊藤 均, Harsojo : 食品照射, 33, 29-32 (1998).
(2000 年 5 月 26 日受理)
|
 |
 |
 |
 |
 |
|
|