恾俆亅侾 兞慄徠幩偵傛傞僕儍僈僀儌乮抝庉乯丄挋憼俉僇寧屻偺忬懺嵍偐傜懳徠嬫丄俁倠俼丆俈倠俼丆侾俀倠俼嬫俈寧俁擔徠幩丄忢壏忢幖壓偵挋憼丄偨偩偟梻侾寧拞弡傛傝俀侽亱俠偺埫強偵挋憼乮弿曽傜丄侾俋俆俋乯
 |
丂壥幚丒偦嵷偼曻幩慄偵懳偡傞姶庴惈偑崅偔丄昳幙偺掅壓傪彽偔偙偲偑懡偄偑丄傾儊儕僇傪偼偠傔僇僫僟丄僜楢偦偺懠奺崙偺尋媶幰偺惛椡揑側尋媶偺惉壥偲偟偰丄悢庬椶偺壥幚丒偦嵷偺挋憼偵桳岠側偙偲偑帵偝傟偨丅偡偱偵僇僫僟丄僜楢偱偼侾俋俇侾擭偵丄暷崙偱偼侾俋俇係擭僕儍僈僀儌偺敪夎梷惂偵曻幩慄傪巊梡偡傞偙偲偑嫋壜偝傟偰偍傝丄僇僫僟丄暷崙偱偼杮擭搙嶻僕儍僈僀儌傛傝徠幩張棟傪婇嬈壔偡傞偨傔柉娫悢幮偑寁夋拞偲偄傢傟傞丅偦偺懠偺壥幚偦嵷偵偮偄偰傕幚梡壔偑恑傔傜傟偰偄傞偑丄尰嵼傾儊儕僇偵偍偄偰嫋壜怽惪偝傟偰偄傞徠幩壥幚偦嵷偺昳栚偼昞俆亅侾偺傛偆偱偁傞丅
暷崙俙俤俠乮尨巕椡埾堳夛乯偺掅慄検徠幩偵傛傞怘昳挋憼偺寁夋乮侾俋俇俀乯傪傒傞偲丄壥幚丒偦嵷偱偼僀僠僑丄儌儌丄僇儞僉僣丄僽僪僂丄僩儅僩側偳偑徠幩偺懳徾偲偟偰宱嵪揑偵丄偐偮彜昳忋偐傜傕桳朷偲偟丄崱屻偼偄傢備傞壥幚偺昞柺嶦嬠偵偮偄偰尋媶傪恑傔傞昁梫惈傪帵嵈偟偨丅僇儕僼僅儖僯傾戝妛偺俵倎倶倝倕傜偼摨埾堳夛偺墖彆偺壓偵偙偺傛偆側壥幚偵偮偄偰尋媶傪峴側偄丄婛偵偦偺惉壥偑曬崘偝傟偰偄傞丅
丂偄傑壥幚丒偦嵷偺挋憼偵懳偡傞曻幩慄棙梡応柺傪弎傋傞偵摉傝丄昅幰偼昞俆亅俀偺傛偆偵暘椶惍棟偟丄偦傟偧傟偺棙梡応柺偵偮偄偰専摙偟偰傒偨偄丅
| 丂昳丂丂丂柤丂 |
嫋壜怽惪 |
丂慄丂検丂俵倰倎倓丂 |
旛丂丂峫 |
| 僕儍僈丂僀丂儌 僞丂儅丂僱丂僊 僀丂丂僠丂丂僑 僩丂丂儅丂丂僩 僇儞僉僣丂丂椶 僒僩僂捫偗壥幚 儌丂丂丂丂丂儌 僯丂儞丂僕丂儞 丂丂丂丂丂丂丂 |
侾俋俇係 侾俋俇係 侾俋俇俇 侾俋俇俈 侾俋俇俁 侾俋俇係 侾俋俇係 侾俋俇係 丂丂丂丂 |
侽丏侽侽俆乣侽丏侽侾 丂丂丂丂丂丂丂丂丂丂 丂丂侽丏俀丂偖傜偄丂 丂丂丂丂丂丂丂丂丂丂 侽丏侽俈俆乣侽丏俆丂 丂丂侽丏俁丂偖傜偄丂 丂丂丂丂丂丂丂丂丂丂 丂丂侽丏侽侾俀丂丂丂 丂丂係丏俆丂丂丂丂丂 |
敪夎梷惂 敪夎梷惂 昞柺嶦嬠 丂丂丂丂 昞柺嶦嬠 丂丂丂丂 丂丂丂丂 敪夎梷惂 嶦丂丂嬠 |
| 丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂乮攡揷丄侾俋俇俆乯 |
| 棙丂梡丂応丂柺 |
丂丂丂揔丂丂梡丂丂慄丂丂検丂丂丂 |
丂丂丂丂丂丂丂丂丂揔丂丂丂丂丂梡丂丂丂丂丂斖丂丂丂丂丂埻丂丂丂丂丂丂丂丂丂 |
|
| 敪丂夎丂梷丂惂 |
侽丏俆乣俆丏侽亊侾侽丒俤乮係乯俼 |
僕儍僈僀儌丄僞儅僱僊丄僯儞僕儞丄僋儕壥幚偺敪崻丄敪夎丄拪戂杊巭側偳丂丂丂丂 |
|
| 弉搙挷惍偦偺懠 丂丂丂丂丂丂丂 嶦丂拵丒嶦丂棏 |
侽丏俆乣俆丏侽亊侾侽丒俤乮俆乯俼 丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂 侽丏侾乣侾丏侽亊侾侽丒俤乮俆乯俼 |
僇僉壥幚偺扙廰丄壥幚偺捛弉懀恑傑偨偼梷惂丄傾僗僷儔僈僗偺慻怐偺擃壔丄儕儞僑 偺儎働昦杊巭丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂 姡憞壥幚丒偦嵷偺嶦拵丄嶦棏丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂 |
|
| 椻 嶦 嬠 |
丂昞柺嶦嬠 丂丂丂丂丂 丂姰慡嶦嬠 |
侾丏侽乣俆丏侽亊侾侽丒俤乮俆乯俼 丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂 侾丏侽乣俆丏侽亊侾侽丒俤乮俇乯俼 |
惗慛壥幚丒偦嵷偺昞柺嶦嬠偵傛傞堦帪揑曐懚丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂 丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂 壥廯丄娛媗椶偺嶦嬠丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂 |
丂偝偒偵弎傋偨傛偆偵傾儊儕僇丄僇僫僟偱偼偡偱偵幚梡揑抜奒偵偒偰偄傞傕偺偱丄傢偑崙偱傕徠幩怘昳偲偟偰嵟傕憗偔巗応偵搊応偡傞傕偺偲婜懸偝傟傞丅
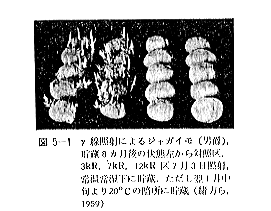
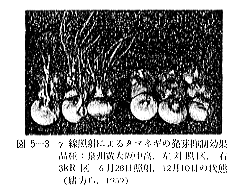
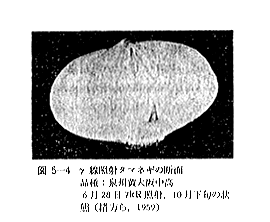
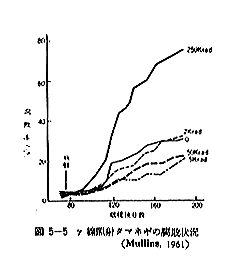
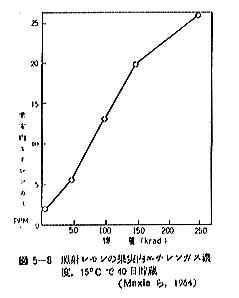
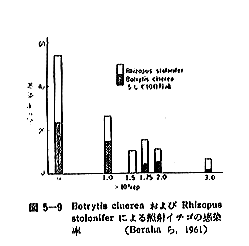
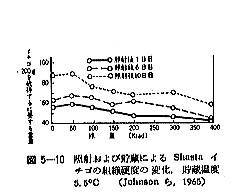
丂僕儍僈僀儌偺敪夎梷惂偵昁梫側慄検偼侾侽倠倰倎倓慜屻偺斾妑揑掅慄検偱懌傝傞偑丄徠幩忦審傗僀儌帺懱偺忬懺側偳偵傛傝偄偔傇傫堎側偭偰偔傞丅恾俆亅侾偼昅幰傜乮侾俋俆俋乯偑媥柊婜偺僕儍僈僀儌乽抝庉乿偵徠幩偟丄挋憼俉僇寧屻偺忬懺傪帵偟偨傕偺偱偁傞偑丄俁倠俼偱偼岠壥偼側偔丄俈倠俼側偄偟侾俀倠俼仏偱岠壥偑姰慡偵傒傜傟傞丅傑偨昳庬乽傾儕乿偺媥柊婜偵徠幩偟偨応崌偱傕俈倠俼偱岠壥偼姰慡偱偁傞丅俽倫倎倰倰倧倵傜乮侾俋俆係乯丄俛倰倧倵値倕倢倢傜乮侾俋俆俈乯丄俹倕倓倕倰倱倧値傜乮侾俋俆俇乯傕戝懱偙偺慄検偱岠壥偺偁傞偙偲傪曬崘偟丄傑偨僇僫僟偵偍偗傞幚尡寢壥乮侾俋俇侾乯偱傕俉倠俼偺慄検偱侾侽昳庬偺僕儍僈僀儌偑偄偢傟傕廫暘偵敪夎偑梷惂偝傟傞偲偟偰偄傞丅偟偐偟昅幰傜乮侾俋俆俋乯偼媥柊婜傪偡偓傞媥柊妎偣偄婜偵徠幩偡傞偲丄俈倠俼偱偼晄廫暘偱挋憼拞敪夎偡傞傕偺傕偁傞偙偲傪抦偭偨丅傑偨敪夎梷惂岠壥偼摨堦慄検偱傕慄検棪偺憡堘偵傛偭偰偐側傝堎側傝丄慄検棪偺崅偄曽偑傛傝岠壥揑偱偁傞丅昞俆乕俁偼乽傾儕乿偵偮偄偰偺幚尡椺偱偁傞偑丄摨偠俀侽倠俼偱傕慄検棪偑掅偄偲挋憼拞偵敪夎偡傞丅偙偺揰俵倎倲倛倳倰乮侾俋俇俁乯傕摨條側偙偲傪巜揈偟偰偄傞丅偟偨偑偭偰僕儍僈僀儌偺応崌丄廂妌屻側傞傋偔憗偔偄傢備傞媥柊婜偵丄崅慄検棪偱徠幩偡傞偙偲偑揔愗偱偁傞偲峫偊傜傟傞丅
丂偝偰偙偺傛偆偵徠幩偟偰敪夎梷惂偝傟偨僕儍僈僀儌偑壔妛惉暘偺忋偵偳偺傛偆側塭嬁傪庴偗傞偐偼嫽枴偁傞栤戣偱偁傞偑丄堦斒揑偵傒偰敪夎梷惂偵懌傞慄検偺斖埻撪偱偼壔妛揑惉暘偵偝傎偳戝偟偨塭嬁偼側偄傛偆偱偁傞丅摐娷検偼恾俆亅俀偺傛偆偵丄徠幩屻忢壏壓偵挋憼偡傞偲徠幩捈屻堦帪揑偵憹壛偟丄偦偺屻尭彮偟偰柍徠幩偺傕偺偵嬤偢偒侾侽寧埲崀嵞傃媫憹偡傞丅偙傟偼偙偺帪婜偵掅壏偵傛偭偰僀儌拞偺僨儞僾儞偑摐偵堏峴偡傞偑丄偦傟偑柍徠幩偺傕偺偱偼敪夎偺偨傔梡偄傜傟傞偺偵懳偟偰丄徠幩偟偨傕偺偱偼敪夎偟側偄偐傜偦偺傑傑拁愊偝傟傞傕偺偲巚傢傟傞丅傾僗僐儖價儞巁偼徠幩偵傛傞捈愙揑塭嬁偼側偄傛偆偱偁傞偑丄俽們倛倰倕倝倐倕倰傜乮侾俋俆俉乯偵傛傞偲挋憼拞偵堦偨傫惗崌惉偝傟憹壛偟偨傾僗僐儖價儞巁偺懝幐偼徠幩僀儌偺曽偑彮側偄偲偄偆丅
丂懠曽徠幩僀儌偺晠攕偼堦斒偵敪夎梷惂偵懌傞慄検偱偼倂倧倰倠倣倎値傜乮侾俋俇侽乯偺帵偡傛偆偵忈奞傪惗偢傞偙偲偼側偄偑丄慄検偑崅偔側傞偲庬乆忈奞偑惗偠丄昦尨嬠偵懳偡傞掞峈惈偑尭彮偟晠攕偡傞偙偲傕抦傜傟偰偄傞丅俛倰倧倵値倕倢倢傜乮侾俋俆俈乯偼侾俋乣俀侽倠倰倕倫乮儗僢僾乯偱徠幩偟偨僀儌偼晠攕偑懡偔側傞偲弎傋丄偦偺尨場偼徠幩偝傟偨僀儌偺廃旂偺宍惉偑慾奞偝傟丄傑偨栘幙壔偑抶墑偝傟傞偺偱徠幩僀儌偑懝彎傪庴偗傞偲偦偙偐傜嬠偑怤擖偟傗偡偔側傝晠攕偑懡偔側傞偲偟偰偄傞丅傑偨俢倳値們倎値傜乮侾俋俆俋乯偼侾俆乣俀俆倠倰倕倫偺慄検偱寬慡側僕儍僈僀儌側傜偽晠攕偼彮側偄偑丄懝彎傪梌偊傞偲偐丄嬠傪愛庢偡傞偲摨條張棟偟偨柍張棟偺傕偺傛傝晠攕偑懡偔側傞偲偄偆丅偟偨偑偭偰徠幩僀儌偼桝憲丄挋憼拞偺庢傝埖偄偵拲堄偟丄側傞傋偔懝彎傪梌偊側偄偙偲偑戝愗偱偁傞丅側偍丄惗棟昦偱偁傞俛倢倎們倠丂倛倕倎倰倲丂偼徠幩偵傛偭偰彮側偔偡傞偙偲偑偱偒傞偲偄傢傟傞乮俵倳倠們倛倝値丆侾俋俇俀乯丅
丂徠幩僀儌偺廳検尭乮儊儀儕乯偺曄壔偼敪夎偑梷惂偝傟傞偨傔柍張棟僀儌偵斾傋偰彮側偄偑丄挿婜忢壏壓偵曻抲偡傞偲乽僔儚乿偑尰傢傟傞偺偱丄僀儌偐傜忲嶶傪杊偖曽朄傪暪梡偡傞偙偲偑昁梫偱偁傞丅偟偐偟側傫傜偐偺曽朄偱忲嶶傪梷惂偡傞偙偲偑偱偒偰傕丄夁幖偵側傞偲晠攕偟傗偡偄偺偱偙偺揰拲堄傪梫偡傞丅偨偲偊偽俙倱倱倕倢倢倐倕倰倗倱傜乮侾俋俇侽乯偼徠幩僀儌傪幒壏壓偱挋憼偡傞偨傔偵徠幩僀儌傪杊陘嵻偱張棟偟偨屻億儕僄僠儗儞偺傛偆側僾儔僗僠僢僋僗僼傿儖儉偵晻擖偟偦偺岠壥傪挷嵏偟偨偑丄悢擔偱柍張棟丄徠幩僀儌偲傕偵怓崄偵曄壔偑婲傞偺偱偙偺傛偆側挋憼朄偑晄揔摉偱偁傞偙偲傪帵偟偰偄傞丅
丂徠幩偵傛傝敪夎偝傟偨僕儍僈僀儌偺歯岲惈偼昞俆乕係偵傒傜傟傞傛偆偵丄俆乣俉侽倠俼偺掱搙偺徠幩偱偼寢壥偼椙岲偱偁傞丅傑偨慻怐偺峝偝偼昳幙傪嵍塃偡傞廳梫側堦場偱偁傞偑丄俆乣俀侽倠俼偺徠幩僀儌偼俆乣俈亷偱侾俉僇寧娫挋憼偟偰傕峝偄忬懺偱曐帩偟偆傞偑丄偦傟埲壓偍傛傃偦傟埲忋偺慄検偱偼廮傜偐偔側傞乮俽倫倎倰倰倧倵傜丄侾俋俆係乯丅俵倎倓倱倕値傜乮侾俋俆俋乯偼暔棟揑曽朄偵傛偭偰徠幩僀儌偺峝偝傪應掕偟丄俋丏俁倠倰倎倓偱徠幩僀儌偺愗抐埑偼掅偔廮傜偐偔側偭偨偲偟偰偄傞丅
丂仏丂俼丗儗儞僩僎儞
| 丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂 丂丂慄丂丂丂丂検丂丂丂偍傛傃丂丂丂慄丂丂検丂丂棪丂丂丂丂 丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂 |
丂丂丂丂挷丂丂丂丂嵏丂丂丂丂寧丂丂丂丂擔丂丂丂丂丂 |
|||||
| 俋丏俈 |
侾侽丏侾俇 |
侾侽丏俁侽 |
侾俀丏俈 |
侾丏俀俁 |
||
| 丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂 丂丂丂丂丂侽丂丂丂丂丂丂丂丂丂丂丂丂丂倰丂丂丂丂丂丂丂丂 |
丂丂仏 丂俁丂 |
丂丂丂丂丂 丂俉俆丂丂 |
丂丂丂丂丂 丂侾侽侽丂 |
丂丂丂丂 丂侾侽侽 |
丂丂丂丂 丂侾侽侽 |
|
| 侾丂丂亊侾侽丒俤乮係乯倰 丂丂丂丂丂丂丂丂丂丂丂丂 |
丂丂丂丂俆侽侽丂丂丂丂俼乛倛倰 侾丏俇亊侾侽丒俤乮俆乯俼乛倛倰 |
丂侽丂 丂侽丂 |
丂侾係丂丂 丂丂侽丂丂 |
丂丂俁係丂 丂丂丂侽丂 |
丂丂俇俆 丂丂丂侽 |
丂丂俈侾 丂丂丂侽 |
| 侾丏俆亊侾侽丒俤乮係乯倰 丂丂丂丂丂丂丂丂丂丂丂丂 |
丂丂丂丂俈俆侽丂丂丂丂俼乛倛倰 侾丏俇亊侾侽丒俤乮俆乯俼乛倛倰 |
丂侽丂 丂侽丂 |
丂丂侽丂丂 丂丂侽丂丂 |
丂丂侾侽丂 丂丂丂侽丂 |
丂丂係侾 丂丂丂侽 |
丂丂俆俉 丂丂丂侽 |
| 俀丂丂亊侾侽丒俤乮係乯倰 丂丂丂丂丂丂丂丂丂丂丂丂 |
丂丂丂侾侽侽侽丂丂丂丂俼乛倛倰 侾丏俇亊侾侽丒俤乮俆乯俼乛倛倰 |
丂侽丂 丂侽丂 |
丂丂侽丂丂 丂丂侽丂丂 |
丂丂丂侽丂 丂丂丂侽丂 |
丂丂侾俀 丂丂丂侽 |
丂丂俀侾 丂丂丂侽 |
| 丂丂仏丗丂敪夎棪乮亾乯丂丂昳庬傾儕丄俈寧俀俋擔徠幩乮弿曽丄侾俋俇係乯 丂丂拲丗丂倰丂偼倰倎倓偲傎傏摨偠扨埵 |
| 丂丂丂丂丂 張棟屻擔悢 丂乮寧乯丂 |
丂丂丂丂丂 僥僗僩偟偨 丂恖偺悢丂 |
丂丂丂丂丂丂丂丂丂丂丂丂歯丂岲丂惈丂偁丂傝丂偲丂偟丂偨丂恖丂悢丂丂丂丂丂丂丂丂丂丂丂丂 |
|||||
| 俠倧値倲 |
侾丆俀俆侽俼 |
俆丆侽侽侽俼 |
俀侽丆侽侽侽俼 |
俉侽丆侽侽侽俼 |
侾侽俇丆俀俆侽俼 |
||
| 丂丂侾丂丂 丂丂係丂丂 丂丂俉丂丂 丂侾俉丂丂 |
丂丂俋丂丂 丂丂俋丂丂 丂侾侾丂丂 丂丂俋丂丂 |
丂俇丂丂 丂俉丂丂 丂係丂丂 丂丂丂丂 |
丂丂俉丂丂丂 丂丂俉丂丂丂 丂侾侽丂丂丂 丂丂丂丂丂丂 |
丂丂俈丂丂丂 丂丂俋丂丂丂 丂丂俈丂丂丂 丂丂俈丂丂丂 |
丂丂丂俇丂丂丂 丂丂丂俉丂丂丂 丂丂侾侽丂丂丂 丂丂丂俉丂丂丂 |
丂丂丂俈丂丂丂 丂丂丂俉丂丂丂 丂丂丂俈丂丂丂 丂丂丂俉丂丂丂 |
丂丂丂丂丂丂丂丂 丂丂丂丂丂丂丂丂 丂丂丂俈丂丂丂丂 丂丂丂丂丂丂丂丂 |
| 丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂乮俽倫倎倰倰倧倵傜丄侾俋俆係乯 丂丂仏丗慄検俼乮儗儞僩僎儞丄侾俼偼栺侾倰倎倓乯 |
|
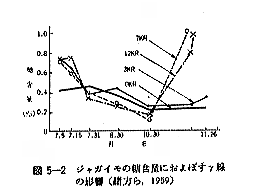 |
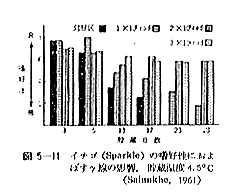
丂僞儅僱僊傕僕儍僈僀儌偲摨偠傛偆偵斾妑揑掅慄検偱敪夎梷惂岠壥偺傒傜傟傞傕偺偱偁傞丅偟偐偟偦偺昁梫慄検偼張棟忦審偵傛偭偰偐側傝嵎偺偁傞偙偲偑曬崘偝傟偰偄傞丅昅幰傜乮侾俋俆俋乯偑僕儍僈僀儌偲摨條偵乽愹廈墿乿僞儅僱僊偵偮偒媥柊婜偍傛傃媥柊妎偣偄婜偺椉帪婜偵偦傟偧傟丄俁丄俈偍傛傃侾俀倠俼傪徠幩偟偨偲偙傠丄徠幩帪婜傪栤傢偢丄傑偨慄検偵娭學側偔偄偢傟傕敪夎偼姰慡偵梷惂偝傟偨乮恾俆乕俁乯丅彫搰傜乮侾俋俇俁乯偼廂妌屻俉俇擔傑偱偵徠幩偡傟偽俁倠俼偱姰慡偵敪夎梷惂偱偒傞偑丄廂妌屻侾侾係擔偱偼侾俆倠俼偱傕岠壥偺側偄偙偲傪曬偠偰偄傞乮昞俆乕俆乯丅偙偺傛偆側廂妌屻偺擔悢偲慄検偲偺娭學偵偮偄偰偼俵倳倢倢倝値倱傜乮侾俋俇侾乯傕摨條側孹岦傪曬崘偟偰偄傞丅
丂偲偙傠偱敪夎梷惂偝傟偨僞儅僱僊偺撪晹偼恾俆乕係偵傒傜傟傞傛偆偵撪夎偺晹暘偩偗偑忈奞傪婲偟妼曄屚巰偡傞丅偙偺撪夎偺妼曄尰徾偼俢倎倢倢倷値傜乮侾俋俆俋乯丒俶倳倲倲倎倢倢傜乮侾俋俇侾乯偵傛偭偰傕娤嶡偝傟偰偄傞偑丄昅幰傜偵傛傟偽撪夎偑忈奞傪婲偡帪婜偼媥柊婜偵徠幩偟偨傕偺傛傝媥柊妎偣偄婜偵徠幩偟偨傕偺偺曽偑傛傝憗偔丄偟偐傕忈奞晹暘偑傛傝戝偒偔側傞丅偟偨偑偭偰僞儅僱僊偺応崌偙偺揰昳幙曐帩忋懡彮栤戣傕偁傞偑丄僕儍僈僀儌摨條撪夎偺傑偩摦偒巒傔側偄帪婜偡側傢偪媥柊婜乕廂妌屻側傞傋偔憗偔徠幩張棟偡傞偙偲偑揔愗偲峫偊傞丅丂昞俆乕俇偼敪夎梷惂偵懌傞昁梫慄検偺尷奅偲昳庬娫偺嵎堎偵偮偄偰傒偨傕偺偱偁傞偑丄偳偺昳庬偲傕俀倠俼偱廫暘梷惂偝傟偰偄傞丅斞捤傜乮侾俋俇俀乯偼戝検張棟偺応崌偼偙傟傛傝傕崅偄慄検偺昁梫側偙偲傪曬崘偟偰偄傞丅傑偨徠幩僞儅僱僊偺敪夎棪偼媴偺戝彫丄挋憼壏搙偵傕塭嬁偝傟傞偲偄傢傟丄媴偺彫偝偄傕偺偺曽偑戝偒偄傕偺傛傝敪夎棪偑戝偱偁傝丄挋憼壏搙偼忢壏乮侾俆亷乣俁侽亷乯傛傝侾俁亷偺曽偑敪夎棪偼戝偱偁傞偲偄偆丅
丂徠幩僞儅僱僊偺廳検尭偺曄壔偼媴偺敪夎偵傛傞懝幐偼彮側偄偑丄俆僇寧娫忢壏曻抲偱偼俀俆亾慜屻尭彮偡傞乮彫搰傜丄侾俋俇俁乯丅偙偺揰偝傜偵廳検偺懝幐傪杊偖曽朄傪岺晇偡傞昁梫偑偁傠偆丅徠幩僞儅僱僊偺晠攕棪偼慄検偑崅偔側傞偲崅偔側傞乮昞俆乕俆丄恾俆乕俆乯丅側偍壔妛揑惉暘偵懳偡傞塭嬁偼傎偲傫偳側偄丅偟偐偟歯岲惈偐傜傒傞偲丄俫倎値値倎値乮侾俋俆俇乯丆俢倎倢倢倷値傜乮侾俋俆俋乯丄俶倳倲倲倎倢倢傜乮侾俋俇侾乯丄俴倕倵倝倱倱傜乮侾俋俇俁乯偺寢壥偐傜傒傞偲丄怓傗慻怐偺峝偝偵偼曄壔偼擣傔傜傟側偄偑丄偄傢備傞僞儅僱僊摿桳偺巋寖枴偑偄偔傜偐敄傟傞傛偆偱偁傞丅
| 徠丂丂幩丂丂擔 丂丂丂丂丂丂丂 乮廂妌屻擔悢乯 |
慄丂丂検 丂丂丂丂 乮倠倰乯 |
丂丂丂 媴丂悢 丂丂丂 |
丂丂丂 晠攕悢 丂丂丂 |
丂丂丂丂敪丂夎丂媴丂悢丂丂丂丂 |
丂丂丂丂丂 慡敪夎媴悢 丂丂丂丂丂 |
丂丂丂丂丂 敪丂夎丂棪 丂丂丂丂丂 |
|
| 挿偝亜侾侽們倣 |
挿偝亜侾侽們倣 |
||||||
| 俠倧値倲倰倧倢 |
丂丂侽丂 |
侾俆俉 |
丂侾丂 |
丂丂俈俁丂丂丂 |
丂丂係俀丂丂丂 |
丂侾侾俆丂 |
丂俈俀丏俉 |
| 丂丂丂丂丂丂丂 丂俉寧俀俋擔丂 丂丂丂丂丂丂丂 丂乮俉俇擔乯丂 丂丂丂丂丂丂丂 |
丂丂俁丂 丂丂俇丂 丂丂俋丂 丂侾俀丂 丂侾俆丂 |
丂係俉 丂係俉 丂係俉 丂係俉 丂係俉 |
丂俆丂 丂侾丂 丂係丂 丂侾丂 丂俀丂 |
丂丂丂侽丂丂丂 丂丂丂侽丂丂丂 丂丂丂侽丂丂丂 丂丂丂侽丂丂丂 丂丂丂侽丂丂丂 |
丂丂丂侽丂丂丂 丂丂丂侽丂丂丂 丂丂丂侽丂丂丂 丂丂丂侽丂丂丂 丂丂丂侽丂丂丂 |
丂丂侽丂丂 丂丂侽丂丂 丂丂侽丂丂 丂丂侽丂丂 丂丂侽丂丂 |
丂丂侽丂丂 丂丂侽丂丂 丂丂侽丂丂 丂丂侽丂丂 丂丂侽丂丂 |
| 丂丂丂丂丂丂丂 丂俋寧俀俇擔丂 丂丂丂丂丂丂丂 丂乮侾侾係擔乯 丂丂丂丂丂丂丂 |
丂丂俁丂 丂丂俇丂 丂丂俋丂 丂侾俀丂 丂侾俆丂 |
丂係俉 丂係俉 丂係俉 丂係俉 丂係俉 |
丂侽丂 丂侽丂 丂侽丂 丂侾丂 丂侽丂 |
丂丂俀侽丂丂丂 丂丂俀俀丂丂丂 丂丂俀俁丂丂丂 丂丂俀係丂丂丂 丂丂俀俈丂丂丂 |
丂丂俀俀丂丂丂 丂丂侾俆丂丂丂 丂丂俀侾丂丂丂 丂丂侾俈丂丂丂 丂丂侾俈丂丂丂 |
丂係俀丂丂 丂俁俈丂丂 丂係係丂丂 丂係侾丂丂 丂係係丂丂 |
丂俉俈丏俆 丂俈俈丏侾 丂俋侾丏俈 丂俉俆丏係 丂俋侾丏俈 |
| 丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂乮彫搰傜丄侾俋俇俁乯 |
| 昳丂丂庬 |
丂丂慄丂丂丂検丂丂丂 |
侾侽寧俀俈擔 |
侾俀寧俀侾擔 |
| 奓捤憗惗 丂丂丂丂 丂丂丂丂 丂丂丂丂 |
丂丂丂丂侽丂丂丂丂俼 侾亊侾侽丒俤乮俁乯俼 俀亊侾侽丒俤乮俁乯俼 俁亊侾侽丒俤乮俁乯俼 |
丂丂俉侽仏丂 丂丂俁侽丂丂 丂丂丂侽丂丂 丂丂丂侽丂丂 |
丂侾侽侽丂丂 丂丂俉侽丂丂 丂丂丂侽丂丂 丂丂丂侽丂丂 |
| 戝嶃拞崅 丂丂丂丂 丂丂丂丂 丂丂丂丂 |
丂丂丂丂侽丂丂丂丂俼 侾亊侾侽丒俤乮俁乯俼 俀亊侾侽丒俤乮俁乯俼 俁亊侾侽丒俤乮俁乯俼 |
丂丂俁侽丂丂 丂丂俁侽丂丂 丂丂丂侽丂丂 丂丂丂侽丂丂 |
丂丂俋侽丂丂 丂丂俉侽丂丂 丂丂丂侽丂丂 丂丂丂侽丂丂 |
| 戝嶃娵丂 丂丂丂丂 丂丂丂丂 丂丂丂丂 |
丂丂丂丂侽丂丂丂丂俼 侾亊侾侽丒俤乮俁乯俼 俀亊侾侽丒俤乮俁乯俼 俁亊侾侽丒俤乮俁乯俼 |
丂丂侾侽丂丂 丂丂丂侽丂丂 丂丂丂侽丂丂 丂丂丂侽丂丂 |
丂丂係侽丂丂 丂丂係侽丂丂 丂丂丂侽丂丂 丂丂丂侽丂丂 |
| 丂仏丗丂敪夎棪乮亾乯丂丂丂丂丂丂丂丂丂丂丂丂乮弿曽丄侾俋俇俁乯 |
 |
 |
 |
丂僯儞僕儞偺拪戂傪杊巭偡傞偺偵傕兞慄徠幩偑桳岠偱偁傞丅俵倎倓倱倕値傜乮侾俋俆俋乯偺寢壥乮昞俆乕俈乯偱偼丄俋丏俁倠倰倎倓偱偐側傝敪夎偼杊巭偱偒傞偑姰慡偱側偔丄挋憼壏搙俀侾亷偱偼俁俈丏俀倠倰倎倓偱丄侾侽亷壓偱偼俋丏俁倠倰倎倓偱姰慡偵敪夎偑梷惂偝傟傞丅傑偨徠幩屻俁僇寧挋憼偟偨応崌偺昳幙偼徠幩偟偨傕偺偺曽偑傛偐偭偨偲偟偰偄傞丅
| 丂慄丂丂検丂 丂丂丂丂丂丂 乮倠倰倎倓乯 |
丂丂丂俀侾亷挋憼丂丂丂丂丂丂丂丂丂丂侾侽亷挋憼丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂 敪夎偺暯嬒怢挿丂乮僀儞僠乯丂丂敪夎偺暯嬒怢挿丂乮僀儞僠乯丂丂丂夎偺廳偝丂丂丂廳検尭 俀廡娫丂丂丂係丂丂丂丂俇丂丂丂丂侾丂丂丂丂俀丂丂丂丂俁丂丂丂丂丂乮亾乯丂丂丂乮亾乯 |
| 丂丂丂丂侽丂 丂丂俋丏俁丂 丂侾俉丏俇丂 丂俁俈丏俀丂 丂俇俆丏侾丂 丂俋俁丏侽丂 |
侽丏俈丂丂俁丏侾丂丂係丏俉丂丂侽丏俈丂丂俆丏俁丂丂俋丏俀丂丂丂侾俇丏係丂丂丂俋丏俀 侽丏俁丂丂侾丏侾丂丂俀丏侽丂丂侽丏俀丂丂侽丏俈丂丂侾丏俈丂丂丂丂俀丏侽丂丂丂侽丏侽 侽丏侾丂丂侽丏俀丂丂侽丏俁丂丂侽丏侾丂丂侽丏俀丂丂侽丏俀丂丂丂丂侽丏侾丂丂丂侽丏侽 侽丏侽丂丂侽丏侽丂丂侽丏侽丂丂侽丏侽丂丂侽丏侾丂丂侽丏侾丂丂丂丂侽丏侽丂丂丂侽丏侽 侽丏侽丂丂侽丏侽丂丂晠丂攕丂丂侽丏侽丂丂侽丏侾丂丂侽丏侾丂丂丂丂侽丏侽丂丂丂侽丏侽 侽丏侽丂丂侽丏侽丂丂晠丂攕丂丂侽丏侽丂丂侽丏侽丂丂侽丏侽丂丂丂丂侽丏侽丂丂丂侽丏侽 |
| 丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂乮俵倎倓倱倕値傜丄侾俋俆俋乯 |
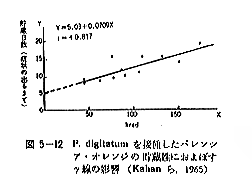
丂僋儕壥幚傕兞慄偺徠幩偵傛傝敪夎敪崻偑梷惂偝傟傞偙偲偑娾揷傜乮侾俋俆俋丄侾俋俇侾乯撧椙乮侾俋俇侾乯偵傛偭偰曬崘偝傟偰偄傞丅偙偺応崌偼僕儍僈僀儌傗僞儅僱僊傛傝傕偄偔傇傫崅偄慄検傪昁梫偲偡傞丅昞俆乕俉偼乽嬧婑乿偵偮偄偰偦偺岠壥傪傒偨傕偺偱丄侾俆倠俼埲忋偺慄検偱梷惂岠壥偺挊偟偄偙偲偑擣傔傜傟傞丅偟偐偟丄俁侽乣俇侽倠俼偺僋儕偼柍徠幩偺僋儕偑敪夎傪巒傔傞崰偵側傞偲丄泱幉晹偵曄怓偡傞傕偺偑尰傢傟傞偺偱丄壛岺尨椏偲偟偰偼偁傑傝揔摉偱側偄丅傢偑崙偱偼僋儕壥幚偺兞慄張棟偼桳岠偱偁傞偲峫偊傜傟傞偐傜丄偝傜偵傛偔尋媶偡傞昁梫偑偁傞丅
丂埲忋曻幩慄徠幩偵傛傞敪夎梷惂岠壥偺応柺傪奣娤偟偨偑丄偙偺岠壥偺尰傢傟傞尨場偵偮偄偰偼尰嵼師偺傛偆偵峫偊傜傟偰偄傞丅乮侾乯暘楐慻怐偺妀巁偺尭彮丄乮俀乯妀抈敀偺摍揹揰偑巁惈懁傊堏峴偡傞側偳偺妀抈敀偺幙揑曄壔丄乮俁乯愼怓懱堎忢側偳偱偁傞丅昞俆乕俋偼俼倳倐倝値傜乮侾俋俇侾乯偺僕儍僈僀儌偺夎偺俼俶俙亅俹偺曄壔傪傒偨傕偺偱偁傞丅
| 應丂掕丂擔 |
慄検亊侾侽丒俤乮係乯俼 |
敪夎 |
敪丂崻 |
敪夎敪崻棪 |
晠攕壥 |
寬丂慡丂壥 |
| 丂丂丂丂丂 侾俀寧俀擔 丂丂丂丂丂 丂丂丂丂丂 |
丂丂俠倧値倲倰倧倢丂丂 丂丂丂丂侾丏俆丂丂丂丂 丂丂丂丂俁丂丂丂丂丂丂 丂丂丂丂俇丂丂丂丂丂丂 |
丂丂 丂丂 丂丂 丂丂 |
俀俇亾 丂丂丂 丂丂丂 丂丂丂 |
丂俀俆亾丂 丂丂俁丂丂 丂丂俈丂丂 丂丂丂丂丂 |
丂丂丂 丂丂丂 丂丂丂 丂丂丂 |
丂俈係亾丂 丂俋俈丂丂 丂俋俁丂丂 侾侽侽丂丂 |
| 丂丂丂丂丂 侾寧侾侾擔 丂丂丂丂丂 丂丂丂丂丂 |
丂丂俠倧値倲倰倧倢丂丂 丂丂丂丂侾丏俆丂丂丂丂 丂丂丂丂俁丂丂丂丂丂丂 丂丂丂丂俇丂丂丂丂丂丂 |
俆侾 丂丂 丂丂 丂丂 |
俁俇丂 丂丂丂 丂丂丂 丂丂丂 |
丂俉俈丂丂 丂侾俉丂丂 丂丂俉丂丂 丂丂丂丂丂 |
丂丂俉 丂丂丂 丂俀俀 丂侾俆 |
丂丂俆丂丂 丂俉俀丂丂 丂俈侽丂丂 丂俉俆丂丂 |
| 丂丂丂廂妌侾侽寧俁擔丄徠幩侾侾寧俇擔丂丂丂丂丂丂丂丂丂丂丂丂丂乮娾揷傜丄侾俋俇侾乯 |
| 慄検乮倠倰倎倓乯 |
侾侾丂丂寧 |
丂丂侾丂丂 |
丂丂俁丂丂 |
丂丂俆丂丂 |
| 丂丂丂丂侽丂丂丂 丂丂丂丂俆丂丂丂 丂丂丂丂俉丂丂丂 丂丂丂侾侽丂丂丂 |
侾俁俋丏俈 丂俇侽丏俉 丂俆俈丏侾 丂係俋丏俁 |
侾俀俀丏係 丂俋侾丏係 丂俉俋丏俈 丂俇俉丏俁 |
侾係侽丏侾 丂俋俀丏俇 侾侽俇丏係 丂俋俀丏俉 |
俀俋侾丏俈 侾俈侽丏侾 侾俀俋丏俉 侾俀俋丏俀 |
| 丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂乮俼倳倐倝値傜丄侾俋俇侾乯 |
丂偙偺応柺傕揔梡慄検偑敪夎梷惂慄検偲傎傏摨偠掱搙偺慄検偱峴側傢傟傞偨傔桳朷偱偁傞偲巚傢傟傞偑丄壥幚丒偦嵷傪懳徾偲偟偨応崌偙傟傑偱偺尋媶寢壥偼傎偲傫偳側偄丅拑捒傜乮侾俋俇係乯偺僋儕壥幚傪梡偄偰偺幚尡寢壥偱偼昞俆乕侾侽偺傛偆偵挋憼拞僔僊僝僂儉僔偺敪惗棪偑戝偱偁偭偨偑丄偦偺敪惗棪偼徠幩偵傛傝傑偨慄検偑崅偔側傞傎偳彮側偔側傞丅偙偺幚尡偱僋儕傪億儕僄僠儗儞戃乮岤偝侽丏侽俁倣倣乯偵晻擖偟偰忢壏壓偵偍偄偨傕偺偼丄悢旵偱傕梒拵偑僋儕壥幚偐傜偱傞偲丄昁偢億儕僄僠儗儞戃傪怘奞偡傞偨傔丄壥幚偺廳検尭偑挊偟偔昳幙偑掅壓偡傞丅偦傟屘徠幩偵傛傝廫暘嶦拵偺栚揑傪払偣傜傟側偄応崌傪峫椂偟丄巆懚偡傞奞拵偵怘奞偝傟側偄傛偆側曪憰傪峴側偆昁梫偑偁傞丅側偍丄姡憞壥幚丄偦嵷偺嶦拵嶦棏傪峴側偄挋憼惈傪崅傔傛偆偲偡傞帋傒傕傒傜傟丄崱屻専摙偡傋偒壽戣偱偁傞丅
| 丂丂丂丂丂丂丂丂 丂慡丂慄丂検丂丂 亊侾侽丒俤乮係乯 丂丂丂丂丂丂丂丂 丂丂丂丂丂丂丂丂 |
忢壏徠幩 俀擔栚丂 丂丂丂丂 丂丂丂丂 丂俽俵丂 |
丂丂丂丂丂丂丂丂丂丂丂丂俁俀丂丂丂擔丂丂丂丂屻丂丂丂丂丂丂丂丂丂丂丂丂丂 |
|||
| 忢壏徠幩忢壏挋憼 丂丂丂丂丂丂丂丂 丂俽丂丂丂俵丂丂 |
椻壏徠幩忢壏挋憼 丂丂丂丂丂丂丂丂 丂俽丂丂丂俵丂丂 |
椻壏徠幩丄椻壏侾俀 擔挋憼屻忢壏挋憼丂 丂俽丂丂丂丂俵丂丂 |
忢壏侾係擔娫妘俀 夞徠幩忢壏挋憼丂 丂俽丂丂丂俵丂丂 |
||
| 丂丂丂丂丂丂丂丂 丂丂丂丂侽丂丂丂 丂丂侾丏俆丂丂丂 丂丂俁丏侽丂丂丂 丂丂俇丏侽丂丂丂 |
丂丂丂倎 丂丂俀丂 丂丂侾丂 丂丂侾丂 丂丂俀丂 |
丂丂丂丂丂丂丂倐 侾俁丂丂俆乮係乯 丂侽丂丂俁丂丂丂 丂侽丂丂侽丂丂丂 丂侽丂丂侽丂丂丂 |
丂丂丂丂丂丂丂丂 侾俀丂丂丂侾丂丂 丂侽丂丂丂係丂丂 丂侽丂丂丂侽丂丂 丂侽丂丂丂侽丂丂 |
丂丂丂丂丂丂丂丂丂 丂俉丂丂丂俇乮俁乯 丂俁丂丂丂係丂丂丂 丂侽丂丂丂侽丂丂丂 丂侽丂丂丂侽丂丂丂 |
丂丂丂丂丂丂丂丂 丂丂丂丂丂丂丂丂 丂俀丂丂俀乮侾乯 丂侾丂丂侾丂丂丂 丂侾丂丂俀丂丂丂 |
| 丂丂拲丂丂俽丗僔僊僝僂儉僔丄丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂乮拑捒傜丄侾俋俇係乯 丂丂丂丂丂俵丗僋儕儈僈 丂丂丂丂丂倎丗俽亄俵乮壥幚撪梒拵悢乯乛俇屄懱 丂丂丂丂丂倐丗俽丆俵乮壥幚撪奜偺梒拵悢乯乛俀係屄懱丄扐偟乮丂乯撪偼鍖偺悢 |
丂堦斒偵惵壥暔偵曻幩慄傪徠幩偡傞偲丄慄検偺嵎偵傛偭偰嵎偼偁傞偑丄暋嶨側惗棟揑曄壔傗慻怐偺擃壔偑婲傞丅偙偺尰徾傪棙梡偡傟偽偁傞偄偼弉搙偺挷惍偑偱偒傞偐傕偟傟側偄偲偄偆娤揰偐傜嵟嬤偐側傝偺曬崘偑弌偝傟偰偄傞丅偟偐偟廂妌屻偺壥幚偺惉弉夁掱偵懳偡傞曻幩慄偺塭嬁偵偮偄偰偼俼倧倣倎値倝乮侾俋俇係乯傜偺曬崘偑偁傞偑丄傑偩廫暘柧傜偐偵偝傟偰偄側偄丅
丂廰奰偵侽丏俁乣侽丏俇俵俼傪徠幩偡傞偲擃壔偟扙廰偡傞丅偙偺偙偲偼偡偱偵憗愳傜乮侾俋俇侾乯丄俲倝倲倎倗倎倵倎傜乮侾俋俇係乯傕巜揈偟偰偄傞偑丄挊幰傜乮侾俋俇俀乯偺峴側偭偨乽暯妀側偟乿偵偮偄偰傒傞偲侽丏侽俆俵俼偱偼柍徠幩偺傕偺偲傎偲傫偳曄傢傝側偄偑丄侽丏俆俵俼偵側傞偲壥幚偑擃壔扙廰偡傞丅偟偐偟俆俵俼偺傛偆側崅慄検偱偼徠幩廔椆帪偡偱偵壥幚偼埫怓傪懷傃丄扙廰偝傟偰偄傞偑壥擏偼曵夡忬懺偵偁傞丅懠偺昳庬偱傕乽暯妀側偟乿偲摨條丄戝懱侽丏俁乣侽丏俇俵俼偱擃壔扙廰偡傞丅傕偭偲傕扙廰偵梫偡傞婜娫偼昳庬偁傞偄偼慄検偵傛偭偰堘偄偑偁傞偑丄慄検偑崅偔側傞傎偳壥幚拞偺壜梟惈僞儞僯儞偺娷検偑尭彮偡傞乮昞俆乕侾侾乯丅懠曽傾僙僩傾儖僨僸僪偺娷検偼崅偔側傞丅傑偨壥幚偺扽巁僈僗攔弌検傕憹戝偡傞丅
丂偙偺傛偆偵徠幩偵傛偭偰壥幚偑擃壔扙廰偡傞偲偄偆偙偲偼幚嵺忋偐傜偼偲傕偐偔丄惗棟妛揑偵偒傢傔偰嫽枴怺偄傕偺偑偁傞丅
| 丂丂丂丂丂丂丂丂丂丂 丂丂丂丂丂丂丂丂丂丂 丂丂丂丂丂丂丂丂丂丂 |
丂徠丂丂幩丂丂屻丂丂擔丂丂悢丂 |
||
| 丂侾丂擔丂 |
丂俁丂丂 |
丂俆丂丂 |
|
| 丂丂丂侽丂丂丂丂丂俼 侾亊侾侽丒俤乮俆乯俼 俁亊侾侽丒俤乮俆乯俼 俇亊侾侽丒俤乮俆乯俼 |
侽丏俁俈亾 侽丏俁俇丂 侽丏侾俀丂 侽丏侽俉丂 |
侽丏係俉 侽丏俁俉 侽丏侽俇 侽丏侽侾 |
侽丏俆俈 侽丏侾侾 侽丏侽俀 侽丏侽侾 |
| 丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂乮弿曽傜丄侾俋俇俀乯 |
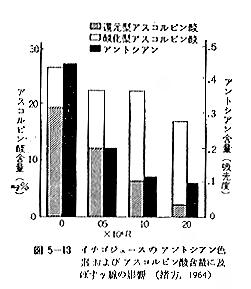
丂僶僫僫偼捛弉壜擻側壥幚偱偁傞偑丄椢怘僶僫僫偵侽丏侾俵俼傪徠幩偡傞偲捛弉偑梷惂偝傟傞偲偄偆俫倎値値倎値乮侾俋俆俇乯偺曬崘偑偁傞丅昅幰傜乮侾俋俇係乯偺幚尡寢壥偐傜傒傞偲丄恾俆乕俇偺傛偆偵侽丏俀俆俵俼偱偼徠幩屻悢擔偱壥旂偑崟曄偡傞丅偟偐偟侽丏侽俆俵俼偱偼柧傜偐偵捛弉偑梷惂偝傟丄柍徠幩偺傕偺偵抶傟偰墿怓傪懷傃傞傛偆偵側傞偑丄偦偺怓偼墭墿怓偱偁傞丅傑偨彮偟弉搙偺恑傫偩傕偺丄偝傜偵墿怓偵側偭偨僶僫僫偵徠幩偟偰傕傗偼傝侽丏俀俆俵俼偱偼壥旂偼崟曄偡傞丅僶僫僫偺捛弉偵敽側偆嵟傕挊偟偄壔妛揑惉暘偺曄摦偼怓丄朏崄偁傞偄偼扽悈壔暔側偳偵傒傜傟傞偑丄偄傑偦偺摐娷検偵偍傛傏偡塭嬁傪傒傞偲昞俆乕侾俀偺傛偆偵側傞丅偡側傢偪椢怓偺僶僫僫偵侽丏侽俆俵俼傪徠幩偡傞偲張棟屻侾俁擔崰偱摐壔偑抶傟傞孹岦偑傒傜傟丄侽丏俀俆俵俼偱偼摐壔偑堦帪懀恑偝傟傞偑丄偦偺屻掆巭偡傞丅
丂偙偺傛偆側偙偲偐傜僶僫僫偼徠幩偵傛偭偰偁傞慄検偱偼柧傜偐偵惗棟揑偵捛弉傪梷惂偡傞偙偲偑偱偒傞偑丄幚嵺忋曻幩慄傪徠幩偡傞偙偲偵傛偭偰僶僫僫傪墿怓壔偝偣丄偁傞偄偼墿怓偵曐帩偝偣傞偙偲偼崲擄偱偁傞偲巚傢傟傞丅
| 丂丂丂丂慄丂丂検丂丂丂丂 丂丂丂丂丂丂丂丂丂丂丂丂 |
丂丂丂丂丂俹倰倕亅俠倢倝倣倎們倲倕倰倝們丂丂丂丂丂丂 侾擔丂丂丂丂係丂丂丂丂侾俁丂丂丂丂侾俇丂丂丂丂侾俋丂 |
丂丂俠倢倝倣倎們倲倕倰倝們丂丂 丂侾丂丂丂丂丂俆丂丂丂丂丂俉丂 |
俹倧倱倲亅俠倢倝倣倎們倲倕倰倝們 丂丂丂侾丂丂丂丂丂丂丂丂係丂丂丂 |
| 丂丂丂丂丂丂丂丂丂丂丂丂 丂丂丂丂侽丂丂丂丂丂丂俼 侽丏俆亊侾侽丒俤乮俆乯俼 俀丏俆亊侾侽丒俤乮俆乯俼 |
丂亾丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂 侽丏俀丂丂侽丏係丂丂侾侾丏侽丂丂侾侽丏侽丂丂丂丂亅丂 侽丏俁丂丂丂亅丂丂丂丂俈丏俉丂丂侾侾丏俆丂丂侾俀丏俁 侽丏俁丂丂俆丏侽丂丂丂係丏俈丂丂丂丂亅丂丂丂丂丂亅丂 |
丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂 侾丏係丂丂侾侾丏侾丂丂侾侾丏俉 侾丏俉丂丂侾俁丏俈丂丂侾侾丏俁 俀丏俀丂丂侾侽丏俉丂丂丂俋丏俉 |
丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂 丂侾俀丏俁丂丂丂丂丂侾俀丏俉丂丂 丂侾俀丏俁丂丂丂丂丂侾俀丏俉丂丂 丂侾侽丏俆丂丂丂丂丂侾俁丏俈丂丂 |
| 丂丂丂丂乮弿曽傜丄侾俋俇係乯 |
 |
丂僩儅僩傕僶僫僫偲摨條偵捛弉壜擻側壥幚偱偁傞偑丄斾妑揑枹弉偺僩儅僩偵侽丏俀乣侽丏係俵俼掱搙偺兞慄傪徠幩偡傞偲拝怓偑抶傟傞丅昞俆乕侾俁偐傜傕傢偐傞傛偆偵徠幩偵傛偭偰僋儘儘僼僀儖偺暘夝偑抶傟丄儕僐僺儞偺敪尰偑梷惂偝傟柧傜偐偵捛弉梷惂偺岠壥偑傒傜傟傞丅偙偺揰俛倳倰値倱丒乮侾俋俆俈乯傗俽倎倢倳値倠倛倕乮侾俋俇侾乯傕弎傋偰偄傞傛偆偵丄偦傟偩偗捛弉婜娫偑墑挿偝傟傞偙偲偵側傞偺偱丄桝憲丄挋憼偵岲搒崌偲側傞偑丄徠幩偵傛傞僩儅僩偺慻怐偺擃壔偼傑偸偑傟偢丄傑偨歯岲惈傕掅壓偡傞孹岦偑傒傜傟傞偺偱乮昞俆乕侾係丄侾俆乯丄側偍堦峫傪梫偡傞偲偙傠偱偁傞丅
| 丂怓丂丂慺丂 |
丂丂慄丂丂丂検丂丂丂 |
丂侽丂丂擔丂 |
丂丂俀丂丂 |
丂丂俈丂丂 |
| 丂丂丂丂丂丂 僋儘儘僼僀儖 丂丂丂丂丂丂 儕僐僺儞丂丂 |
丂丂丂丂侽丂丂丂丂俼 俁亊侾侽丒俤乮俆乯俼 丂丂丂丂侽丂丂丂丂俼 俁亊侾侽丒俤乮俆乯俼 |
侾丏侾俀俆倎 侽丏俋係侽丂 侽丏侽俇俇倐 侽丏侽俉侽丂 |
侽丏俆侽侽 侽丏俇俁侽 侽丏俀俈侽 侽丏侾俀侽 |
侽丏侽侽俆 侽丏俁俈俆 侽丏俇係侽 侾丏侽侽侽 |
| 丂丂丂丂倎丗攇挿俇俇侽倣兪偺媧岝搙 丂丂丂丂倐丗攇挿係俈侽倣兪偺媧岝搙丂丂丂丂丂丂丂乮弿曽傜丄侾俋俇係乯 |
| 慄検乵侾侽丒俤乮俆乯俼乶 |
丂徠幩捈屻丂 |
俉擔栚丂 |
| 丂丂丂丂丂侽丂丂丂丂丂丂 丂丂丂丂丂俀丂丂丂丂丂丂 丂丂丂丂丂係丂丂丂丂丂丂 |
侾丏係俈倠倗 侽丏俇係丂丂 侽丏俆侽丂丂 |
侽丏俉俇 侽丏係俇 侽丏係俈 |
| 丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂乮拑捒傜丄侾俋俇俁乯 |
| 丂丂丂丂丂丂丂丂丂俛倰倕倎倠倕倰丂丂丂丂丂丂丂丂 慄検乵侾侽丒俤乮俆乯俼乶丂丂徠幩屻俉擔栚弴埵崌寁 |
丂丂丂丂丂丂丂俴倝倗倛倲亅倫倝値倠丂丂丂丂丂丂丂 慄検乵侾侽丒俤乮俆乯俼乶丂丂徠幩屻俉擔栚弴埵崌寁 |
| 丂丂丂丂丂侽丂丂丂丂丂丂丂丂丂丂丂丂俀俉丂丂丂丂 丂丂丂丂丂俀丂丂丂丂丂丂丂丂丂丂丂丂俁俇丂丂丂丂 丂丂丂丂丂係丂丂丂丂丂丂丂丂丂丂丂丂俆俀丂丂丂丂 |
丂丂丂丂丂侽丂丂丂丂丂丂丂丂丂丂丂丂俀俋丂丂丂丂 丂丂丂丂丂侽丏俆丂丂丂丂丂丂丂丂丂丂俁侽丂丂丂丂 丂丂丂丂丂俀丏俆丂丂丂丂丂丂丂丂丂丂係俁丂丂丂丂 |
| 丂丂丂俴丏俽丏俢丗侾俀乮俆亾乯丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂乮弿曽傜丄侾俋俇係乯 丂丂丂丂丂丂丂丂丂侾俉乮侾亾乯 |
丂俵倎倶倝倕傜乮侾俋俇係乯偵傛傞偲丄侽丏俀俆俵俼傑偨偼偦傟埲忋偺慄検傪庴偗偨梞僫僔乽僶乕僩儗僢僩乿偼捛弉偑梷惂偝傟傞丅傑偨堦搙弉搙偺恑峴偑梷惂偝傟偨壥幚偼僄僠儗儞僈僗偵懳偡傞斀墳惈偑掅壓偡傞偲偄偆丅偟偐偟拑捒傜乮侾俋俇俁乯偑峴側偭偨寢壥偱傕侽丏俁乣侽丏俇俵俼偺徠幩偱奜娤忋偺梷惂岠壥偼傒傜傟偨偑丄壥幚偼偐側傝擃壔偡傞乮昞俆乕侾俇乯丅偦偺廮傜偐偝偼偪傚偆偳柍張棟壥偺弉壥乮俁擔栚乯偺偦傟偵嬤偄丅偙偺揰弉搙偺憡堘偵傛偭偰惗偢傞梞僫僔壥幚偺峝偝傪曻幩慄徠幩偵傛偭偰嬒堦偵偡傞偙偲偑偱偒傞偐傕偟傟側偄丅
| 慄検乵侾侽丒俤乮俆乯俼乶 |
丂徠幩捈屻丂 |
丂俁擔栚丂 |
丂俈擔栚丂 |
| 丂丂丂丂丂侽丂丂丂丂丂丂 丂丂丂丂丂侾丂丂丂丂丂丂 丂丂丂丂丂俁丂丂丂丂丂丂 丂丂丂丂丂俇丂丂丂丂丂丂 |
丂俁丏侾倠倗 丂俁丏俇丂丂 丂侾丏俇丂丂 丂侽丏俉丂丂 |
丂侾丏俁丂 丂侾丏俈丂 丂侾丏係丂 丂侾丏俀丂 |
丂侽丏俆丂 丂侽丏係丂 丂侽丏俉丂 丂侾丏俀丂 |
| 丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂乮拑捒傜丄侾俋俇俁乯 |
丂椢怓偺儗儌儞偵侽丏侾俵俼傪徠幩偡傞偲椢怓偑柍張棟偺傕偺傛傝憗偔徚幐偡傞丅偟偐偟慄検偑崅偔側傞偲壥幚偼僽儘儞僘怓偵側傞乮俵倎倶倝倕傜丄侾俋俇係乯丅偙偺傛偆側尰徾偼憗惗壏廈偱傕傒傜傟傞乮昅幰傜丄侾俋俇俀乯丅側偍儗儌儞壥幚偺撪晹偼恾俆乕俈偵傒傜傟傞傛偆偵昛偺偆偲昛偺偆偲偺娫偵戝偒側嬻寗偑偱偒傞丅傑偨儗儌儞偼僄僠儗儞僈僗偵傛偭偰捛弉偑懀恑偝傟傞偑丄崅偄慄検傪庴偗偨壥幚傎偳壥幚撪晹偵崅偄擹搙偺僄僠儗儞僈僗偑娷傑傟傞傛偆偵側傝乮恾俆乕俉乯丄傾僗僐儖價儞巁偲僋僄儞巁偑挋憼拞偵尭彮偡傞偙偲偼慄検偑崅偔側傞傎偳戝偱偁傞丅
 |
 |
丂俵們俙倰倓倢倕傜乮侾俋俆俇乯偼儕儞僑偵兞慄傪徠幩偡傞偲壥幚峝搙偑掅壓偟丄偦傟偵敽偭偰晄梟惈儁僋僠儞偑尭彮偡傞偙偲丄傑偨俽倧倣倧倗倷倝傜乮侾俋俇係乯偼僙儖儘乕僗傕尭彮偡傞偙偲傪曬崘偟偰偄傞偑丄嵟嬤俵倎倱倱倕倷傜乮侾俋俇係乯偼侾侽侽倠倰倎倓偺慄検偱挋憼拞偺儎働昦偲俛倰倧倳倵値丂們倧倰倕偺敪惗棪傪彮側偔偡傞壜擻惈偑偁傞偲曬偠偨丅偟偐偟徠堜傜乮侾俋俇俆乯偼峠嬍偱偼俀侽侽倠倰倎倓埲忋丄崙岝偱偼係侽侽倠倰倎倓埲忋偺徠幩傪庴偗傞偲壥幚偵尠挊側忈奞偑尰傢傟傞偲偟偰偄傞偺偱丄曻幩慄徠幩偵傛傞挋憼拞偺惗棟昦敪惗杊巭偵偮偄偰偼側偍傛偔尋媶偡傞昁梫偑偁傞丅傑偨俥倕倰値倎値倓倕倱傜乮侾俋俇俀乯偼徠幩儕儞僑傪俁亷壓偱挋憼偡傞偲徠幩壥幚偺揌掕巁搙偑媫寖偵尭彮偡傞偑丄尭彮偡傞巁偼庡偵儕儞僑巁偱偁傞偲偄偆丅
丂偄偢傟偵偣傛徠幩傪庴偗偨儕儞僑壥幚偼忋婰偺傛偆側庬乆偺惗棟壔妛揑曄壔傪帵偡偙偲偼柧傜偐偱偁傞偑丄墳梡柺偱偼側偍崱屻偺尋媶偵傑偮偲偙傠偑懡偄丅
丂傾僗僷儔僈僗偺梒宻偵兞慄傪徠幩偡傞偲丄昞俆乕侾俈偺傛偆偵慄検偑崅偔側傞偲廮傜偐偝偑憹偡偙偲偑抦傜傟傞丅俽倎倢倳値倠倛倕倰乮侾俋俆俋乯偼偙偺偙偲偐傜娛媗惢憿偺嵺崱傑偱幪偰傜傟偰偄偨傾僗僷儔僈僗偺峝偄晹暘傪棙梡偱偒傞偐傕抦傟側偄偲弎傋偰偄傞丅
| 丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂 慄検棪亊侾侽丒俤乮俇乯倰倎倓乛倛倰 |
丂丂丂丂丂丂慄丂丂丂検丄丂丂侾侽丒俤乮俇乯倰倎倓丂丂丂丂丂 們倧値倲倰倧倢丂丂侽丏俋俁丂丂丂丂俀丏俈俋丂丂丂俆丏俆俉丂 |
|||
| 丂丂丂丂丂丂侽丏侽俋俁丂丂丂丂丂丂 丂丂丂丂丂丂侽丏係俇俆丂丂丂丂丂丂 丂丂丂丂丂丂侽丏俋俁侽丂丂丂丂丂丂 |
丂丂俋侽侽丂丂 丂丂俋侽侽丂丂 丂丂俋侽侽丂丂 |
丂丂俉俆侽丂丂 丂丂俉侽侽丂丂 丂丂俈俋俆丂丂 |
丂丂俇俋俆丂 丂丂俇俉俆丂 丂丂俇俇俆丂 |
丂丂俇俇俆丂 丂丂俇俇侽丂 丂丂俇侾俆丂 |
| 丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂乮俽倎倢倳値倠倛倕傜丄侾俋俆俋乯 |
丂偙傟偼掅慄検偺徠幩偱壥幚丒偦嵷偵晅拝偡傞晠攕嬠傪嶦偟惵壥暔偺堦帪揑曐懚傪寁傠偆偲偡傞傕偺偱偁傞丅懡偔偺応崌掅壏挋憼偲暪梡偡傞偙偲偵傛偭偰傛傝岠壥揑寢壥傪摼傞偙偲偑偱偒傞丅偙偺応柺偱揔梡偝傟傞慄検偼壥幚偦嵷偺弉搙挷惍偵梡偄傜傟傞慄検偲傎傏摨條偱偁傞偨傔丄嶦嬠岠壥偲摨條偵壥幚偦嵷帺恎偺徠幩偵敽偆惗棟壔妛揑曄壔傪丄幚梡壔偵偁偨偭偰廫暘擣幆偟偰偍偔昁梫偑偁傞丅傕偪傠傫偙偺応崌偼徠幩屻嬠偵傛傞擇師姶愼傪庴偗側偄傛偆偵曪憰偡傞昁梫偑偁傞偑丄曪憰梕婍撪偺惵壥暔偑偱偒傞偩偗惓忢側屇媧傪峴側傢偣傞傛偆偵峫偊傞昁梫偑偁傞丅尰嵼偙偺晹栧偼傾儊儕僇側偳偱偼曻幩慄偺岠壥揑棙梡応柺乮昞俆乕侾俉乯偲偟偰幚梡壔偝傟傛偆偲偟偰偄傞偲偙傠偱傕偁傝丄傢偑崙偱傕崱屻偝傜偵尋媶偝傟傞傕偺偲巚傢傟傞丅偄傑悢庬壥幚丒偦嵷偺昦尨嬠巺偵懳偡傞曻幩慄偺抳巰慄検偼丄俛倕倰倎倛倎傜乮侾俋俇侽乯偺挷傋偨寢壥偱偼昞俆乕侾俋偺偲偍傝偲側傞丅埲壓悢庬壥幚偦嵷偵偮偄偰偦偺岠壥傪弎傋傞丅
| 丂昳丂丂丂丂丂丂柤丂 |
丂丂張棟朄丂 |
丂嵟揔慄検丂丂 |
丂挋憼壏搙丂丂 |
丂挋憼擔悢丂丂 |
丂尷奅慄検丂丂 |
| 丂丂丂丂丂丂丂丂丂丂 僆丂丂儗丂丂儞丂丂僕 儗丂丂丂儌丂丂丂丂儞 僀丂丂丂僠丂丂丂丂僑 儌丂丂丂丂丂丂丂丂儌 僽丂丂丂僪丂丂丂丂僂 傾丂僾丂儕丂僐僢僩丂 仏傾僗僷儔僈僗丂丂丂 仏僗僫僢僾丒價乕儞僘 仏壴丂丂儎丂僒丂僀丂 仏僩僂儌儘僐僔丂丂丂 僩丂丂丂儅丂丂丂丂僩 儂僂儗儞僜僂丂丂丂丂 |
丂丂丂丂丂丂 丂丂丂丂丂丂 丂丂丂丂丂丂 丂丂丂丂丂丂 丂丂丂丂丂丂 丂丂丂丂丂丂 丂丂丂丂丂丂 丂丂丂丂丂丂 恀丂丂丂丂嬻 丂丂丂丂丂丂 俶俀僈僗抲姺 丂丂丂丂丂丂 恀丂丂丂丂嬻 |
乮俵倰倎倓乯丂 丂侽丏俀俆丂丂 丂侽丏俀侽丂丂 丂侽丏俀侽丂丂 丂侽丏俁侽丂丂 侽丏侾乣侽丏俀 丂侽丏俀丂丂丂 丂侽丏俆丂丂丂 丂侽丏俈丂丂丂 丂侽丏俈丂丂丂 丂侾丏侽丂丂丂 丂侽丏俀俉丂丂 丂侽丏俋俁丂丂 |
丂丂乮亷乯丂丂 丂丂丂丂丂丂丂 丂丂侾俁丂丂丂 丂丂丂侽丂丂丂 丂丂丂俆丂丂丂 丂丂丂俆丂丂丂 丂丂丂俆丂丂丂 丂丂丂俆丂丂丂 丂丂丂丂丂丂丂 丂丂丂俆丂丂丂 丂丂丂俁丂丂丂 丂俀侾乣俀係丂 丂丂丂俆丂丂丂 |
丂丂丂丂丂丂丂 丂俀乣俁攞丂丂 丂丂侾俈丂丂丂 丂丂侾係丂丂丂 丂丂侾侽丂丂丂 丂丂俀俉丂丂丂 丂丂係俀丂丂丂 丂丂俀侽丂丂丂 丂丂丂俀攞丂丂 丂丂俀俈丂丂丂 丂俁侽侽丂丂丂 丂丂侾侾丂丂丂 丂丂侾俉丂丂丂 |
丂乮俵倰倎倓乯 丂丂侽丏係俆丂 丂丂丂丂丂丂丂 丂丂侽丏俁丂丂 丂丂侽丏俆丂丂 丂丂侽丏俁丂丂 丂丂侽丏係丂丂 丂丂丂丂丂丂丂 丂丂侽丏俋丂丂 丂丂丂丂丂丂丂 丂丂丂丂丂丂丂 丂丂丂丂丂丂丂 丂丂丂丂丂丂丂 |
| 丂丂仏丗搾捠偟張棟屻徠幩偡傞丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂乮攡揷丄侾俋俇俆乯 |
| 丂丂丂丂丂丂丂 丂婑庡偺庬椶丂 |
丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂 丂丂丂丂旝丂丂丂丂惗丂丂丂丂暔丂丂丂丂偺丂丂丂丂庬丂丂丂丂椶丂丂丂丂 |
丂丂丂慄丂検丄侾侽丒俤乮俆乯倰倎倓丂丂丂丂丂 丂丂姦揤攟婎丂丂丂丂丂丂丂丂丂婑庡愙庬丂丂丂 |
| 僽僪僂丒僀僠僑 儗丂丂儌丂丂儞 丂丂丂丂丂丂丂 丂丂丂丂丂丂丂 儌丂丂丂丂丂儌 丂丂丂丂丂丂丂 丂丂丂丂丂丂丂 儕丂丂儞丂丂僑 僕儍僈丂僀丂儌 丂丂丂丂丂丂丂 僞丂儅丂僱丂僊 僩丂丂儅丂丂僩 |
俛倧倲倷倳倲倝倱丂們倝値倕倰倕倎丂俹倕倰倱丏倕倶俥倰丏丂丂丂丂丂丂丂 俹倕値倝們倝倢倢倝倳倣丂倓倝倗倝倲倎倲倳倣丂俽倎們們丏丂丂丂丂丂丂丂 俠倢倎倓倧倱倫倧倰倝倳倣丂倱倫丏丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂 俙倢倲倕倰値倎倰倝倎丂們倝倲倰倝丂俤倢倢丏仌丂俹倝倕倰們倕丏丂丂丂丂 俵倧値倝倢倝値倝倎丂倖倰倳們倲倝們倧倢倎丂乮倂倝値倲丏乯丂俫倧値倕倷 俼倛倝倸倧倫倳倱丂値倝倗倰倝們倎値倱丂俤倛倰倕値倐丏倕倶俥倰丏丂丂丂 俢倝倫倢倧倓倝倎丂値倎倲倎倢倕値倱倝倱丂俹丏俤倴倎値倱丏丂丂丂丂丂丂 俹倕値倝們倝倢倢倝倳倣丂倕倶倫倎値倱倳倣丂乮俴俲丏乯丂俿倛倧倣丂丂丂 俹倛倷倲倧倫倛倲倧倰倎丂乮俵倧値倲乯倓丂俛倷丂丂丂丂丂丂丂丂丂丂丂丂 俹倷倲倝倳倣丂倓倕倐倎倰倷倎値倳倣丂俫倕倱倱倕丂丂丂丂丂丂丂丂丂丂丂 俛倧倲倰倷倲倝倱丂倎倢倢倝倝丂俵倳値値丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂 俙倢倲倕倰値倎倰倝倎丂倲倕値倳倝倱丂俙倳們倲丂丂丂丂丂丂丂丂丂丂丂丂 |
侽丏俋俆乣侾丏俉俇丂丂丂俀丏俈係乣係丏俆俇丂 侾丏侾丂乣侾丏係俉丂丂丂侾丏俉俀乣俀丏侾侽丂 俀丏俆俇乣係丏俀俈丂丂丂係丏俆俇乣俋丏侾俀丂 係丏俀俈乣係丏俆俇丂丂丂俀丏俈係乣係丏俆俇丂 侾丏俁俉乣侾丏俉俆丂丂丂侾丏俁俈乣侾丏俉俀丂 俀丏俈係乣俁丏俆俇丂丂丂侾丏俉俀乣俀丏俀俉丂 係丏俇俆乣俉丏俋侾丂丂丂俀丏俈係乣係丏俆俇丂 侾丏俁俆乣侾丏係侽丂丂丂侾丏俉俀乣俀丏俈係丂 丂丂侽丏俀俆丂丂丂丂丂丂丂丂侽丏俀俆丂丂丂丂 侾丏俉俀乣俀丏俀俉丂丂丂侾丏俆俈乣侾丏俉俀丂 侾丏俁俁乣侾丏俉俁丂丂丂俀丏俈係乣俁丏俇俆丂 係丏俀丂乣係丏俆俈丂丂丂俀丏俈係乣係丏俆俇丂 |
| 丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂乮俛倕倰倎倛倎傜丄侾俋俇侽乯 |
丂俛倕倰倎倛倎傜乮侾俋俆俈乯偵傛偭偰俀侽侽倠倰倕倫偺慄検偱丄忢壏壓偍傛傃椻憼暪梡偵偍偄偰傕僀僠僑偺挋憼婜娫傪墑挿偡傞偙偲偑壜擻偱偁傞偙偲偑曬崘偝傟偰埲棃丄俶倕倢倱倧値傜乮侾俋俆俋乯丄俽倎倢倳値倠倛倕乮侾俋俇侾乯丄斞捤傜乮侾俋俇俀乯丄弿曽傜乮侾俋俇俀乯丄俵倎倶倝倕傜乮侾俋俇俆乯偵傛偭偰曻幩慄徠幩偵傛傞僀僠僑偺挋憼岠壥偑曬偠傜傟偨丅偦偺揔梡慄検偼戝懱俀侽侽倠倰倎倓偱偁傞偑丄偙偺応崌徠幩偵傛偭偰晠攕嬠偑嶦嬠偝傟敪僇價偑杊巭偝傟偰傕僀僠僑偦偺傕偺偺昳幙偑楎壔偡傞傛偆偱偼巚傢偟偔側偄偺偱廫暘拲堄傪梫偡傞丅偄傑俛倕倰倎倛倎傜乮侾俋俇侾乯偵傛傞偲侽丏侾乣侽丏俁俵倰倕倫偱俀侾亷壓偱俁擔娫丄俆亷壓偱侾侽擔娫僇價偺敪惗傪梷偊傞偙偲偑偱偒傞偲偄偆丅恾俆乕俋偼俆亷壓偱侾侽擔娫挋憼偟偨応崌偺柍張棟偲張棟嬫偺俛倧倲倰倷倲倝倱丂們倝値倕倰倕倎丂偲俼倛倝倸倧倫倳倱丂倱倲倧倢倧値倝倖倕倰丂偺姶愼棪傪帵偟偰偄傞丅柧傜偐偵侽丏侾俵倰倕倫埲忋偱姶愼棪偺尭彮偡傞偙偲偑抦傜傟傞丅堦曽恾俆乕侾侽偼俰倧倛値倱倧値傜乮侾俋俇俆乯偺僋儔僢僔儏丒僥僗僩偺寢壥傪帵偟偨傕偺偱偁傞偑丄慄検偑崅偔側傞傎偳壥幚偑擃壔偡傞丅曻幩慄偵傛偭偰怉暔慻怐偺擃壔偺偍偙傞偙偲偼堦斒揑尰徾偱偁傞偑丄僀僠僑偼傕偲傕偲慻怐偺廮傜偐偄壥幚偩偗偵偙偺揰栤戣偑偁傞丅偙偺傛偆側偙偲偐傜俽倎倢倳値倠倛倕乮侾俋俇侾乯偼悢昳庬偺僀僠僑偵偮偄偰昳庬娫偺憡堘傪挷傋丄曻幩慄張棟偵揔偟偨昳庬偺偁傞偙偲傪尒偄偩偟偨丅偦偺偆偪嵟傕椙岲側寢壥傪摼偨昳庬偺歯岲惈僥僗僩偺寢壥傪帵偡偲恾俆乕侾侾偺傛偆偵丄係丏俆亷壓偱俀俉擔挋憼偟偰傕傑偩崅偄歯岲惈偺偁傞偙偲偑傢偐傞丅傑偨俿倰倳倕倢倱倕値乮侾俋俇俁乯偼徠幩僀僠僑偺歯岲惈偵娭偟丄徠幩捈屻徠幩壥幚偺歯岲惈偼偄偔傜偐掅壓偡傞偑挿婜挋憼偡傞偲丄張棟壥幚偺曽偑昳幙忋傛偔側傞丅傑偨摨巵傜偼廂妌擔偺揤岓偺椙斲偑僀僠僑偺挋憼惈偵挊偟偔塭嬁偡傞偙偲傕曬崘偟偰偄傞丅徠幩張棟偵懳偡傞揔惈偼弉搙偵傕娭學偟丄斞捤傜乮侾俋俇俀乯偵傛傞偲姰弉偺傕偺傛傝丄僌儕乕儞僠僢僾偺傕偺偺曽偑傛偄寢壥偑摼傜傟偨偲偄偆丅僀僠僑偺傾儞僩僔傾儞怓慺偍傛傃傾僗僐儖價儞巁偼侽丏俀俆俵俼偱偼斾妑揑塭嬁偼彮側偄偑侽丏俆俵俼偵側傞偲柧傜偐偵尭彮偡傞偲偄偆拑捒傜乮侾俋俇俆乯偺曬崘傕傒傜傟傞丅
丂側偍徠堜傜乮侾俋俇係乯偼俛丏們倝値倕倰倕倎丂俹倕倰倱傪梡偄偨幚尡偱崅慄検棪乮侾俵倰倎倓乛倛倰乯偱徠幩偡傞偲侽丏俉俵倰倎倓偱姰慡偵嬠傪巰柵偡傞偙偲偑偱偒傞偑丄摨慄検偺掅慄検棪乮侽丏侽俀俵倰倎倓乛倛倰乯偱偼岠壥偑尭彮偡傞偲偄偭偰偄傞丅
 |
 |
 |
丂僇儞僉僣椶偵懡偔拝偔儁僯僔儕僂儉嬠偼俛倕倰倎倛倎傜乮侾俋俆俈丄侾俋俆俋乯偵傛傟偽丄曻幩慄偵懳偡傞掞峈惈偑斾妑揑庛偄偺偱丄椙岲側寢壥偑婜懸偝傟傞偑丄壥幚帺懱偺慻怐丄怓偺曄壔偑婲偙傝傗偡偄偲偄傢傟傞丅偟偐偟摨巵傜偺幚尡偱偼僱乕僽儖丒僆儗儞僕偵愛庢偟偨俹倕値倝們倝倢倢倝倳倣丂倝倲倎倢倝們倳倣偼侽丏俀俵倰倎倓偖傜偄偺慄検偱敪惗偑杊巭偝傟丄壥幚偺昳幙傕傎偲傫偳掅壓偣偢丄俀侾亷偱柍張棟偺傕偺偑係擔偟偐曐懚偝傟側偄偺偵張棟偟偨傕偺偱偼俀侽擔丄俆亷偱偼偦傟偧傟侾俆擔丄俇俆擔偲側偭偨偺偱丄徠幩偵傛傞挋憼惈偺憹恑偼廫暘擣傔傜傟偰偄傞丅偝傜偵摨巵傜乮侾俋俇係乯偼慄検棪傪偐偊偰偦偺岠壥傪挷傋丄侽丏侽俀俵倰倎倓乛倣倝値偱偼侽丏侾俉俀俵倰倎倓偱姰慡偵俹丏倝倲倎倢倝們倳倣丂傪巰柵偝偣傞偙偲偑偱偒傞偑丄侽丏侽侽俁俵倰倎倓乛倣倝値偱偼摨慄検偱傕姰慡偱側偄偲偟偨丅傑偨嬤偔俲倎倛倎値傜乮侾俋俇俆乯偼僶儗儞僔傾僆儗儞僕傪梡偄偰徠幩慄検偲挋憼擔悢偲偺娭學傪挷傋恾俆乕侾俀傪摼偰偄傞丅
丂偙偺傛偆側偙偲偐傜傢偑崙偱傕壏廈儈僇儞側偳偵偮偄偰専摙偝傟傟偽偁傞偄偼嫽枴偁傞寢壥偑摼傜傟偆傞偐傕偟傟側偄丅
 |
丂俶倕倢倱倧値傜乮侾俋俆俋乯偵傛傞偲俛丏們倝値倕倰倕倎丂偼侽丏俀俵倰倎倓偱侾係擔娫偙偺嬠偺惗堢傪慾巭偡傞偙偲偑偱偒傞偑丄徠幩忈奞偑戝偒偔丄俵倎倶倝倕傜乮侾俋俇係乯偼徠幩屻偺幭偨傛偆側崄傝偲慻怐偺擃壔偐傜徠幩僽僪僂偑徚旓幰偵庴偗擖傟傜傟傞偐偳偆偐媈傢偟偄偲曬崘偟偰偄傞丅傑偨斞捤傜乮侾俋俇係乯偼曻幩慄張棟偵偁偨偭偰偦偺摿桳偺惈幙傪幐側傢偣偟傔側偄慄検偼侽丏侾俵倰倎倓偱偁傞偲偟偰偄傞丅
丂儌儌偱偼俛倕倰倎倛倎傜乮侾俋俆係乯偼柍張棟壥偼俛倰倧倵値丂倰倧倲偺偨傔俆擔娫偱姰慡偵晠攕偡傞偑丄侽丏俀俵倰倎倓傪徠幩偡傞偲曐懚婜娫傪侾侽擔娫墑挿偡傞偙偲偑偱偒傞偲弎傋偰偄傞丅俢倛倎倢倝倵倧倢傜乮侾俋俇俁乯偵傛傞偲億儕僄僠儗儞曪憰偟侽丏侾乣侽丏俁俵倰倎倓傪徠幩偡傞偲椻憼婜娫傪侾俆乣俀侽擔傑偱墑挿偡傞偙偲偑偱偒傞偲偄偆丅
丂僗僀乕僩丒僠僃儕乕偵偮偄偰偼俠倧倧倫倕倰傜乮侾俋俇俁乯偵傛傝椙岲側寢壥偑曬崘偝傟偰偄傞丅係丏俆亷丄幖搙俉侽亾偱挋憼偟偨応崌丄侽丏係俵倰倎倓埲忋偺慄検偱偐偭曄偡傞偑丄侽丏俁俵倰倎倓偱偼栺俁侽擔挋憼婜娫傪墑挿偡傞偙偲偑壜擻偱偁傞丅偟偐偟捛弉偲怓偺敪尰偑抶傟傞丅傑偨僇僾僞儞偺傛偆側壔妛暔幙偱張棟偟僾儔僗僠僢僋僗僼僀儖儉偱曪憰偟偰徠幩偡傞偲椙偄寢壥偑摼傜傟偨偲偄偆丅偙偺幚尡偱敪惗偟偨旝惗暔偵偮偄偰傒傞偲丄俹倕値倝們倝倢倢倝倳倣嬠傛傝俙倢倲倕倰値倕倰倝倎嬠偺曽偑兞慄偵懳偟偰掞峈惈偑戝偱偁傞偙偲傪帵偟偰偄傞丅
丂埲忋偺傎偐僩儅僩乮俽倎倢倳値倠倛倕丆侾俋俇侾丄俿倰倳倕倢倱倕値丆侾俋俇俁乯丄儕儞僑乮徠堜傜丄侾俋俇俆乯丄梞僫僔乮俛倕倰倎倛倎丆侾俋俇係乯丄僉儍儀僣乮俽倎倢倳値倠倛倕傜丄侾俋俆俋乯丄僗僀乕僩丒僐乕儞乮俽倎倢倳値倠倛倕傜丄侾俋俆俋乯側偳偑偁傞偑丄摿婰偡傞傛偆側椙偄寢壥偼摼傜傟偰偄側偄丅
丂壛岺忋偺嶦嬠偲側傞偲丄姰慡偵嶦嬠偡傞昁梫偑偁傞傢偗偱丄偦偺昁梫慄検偼侾俵倰倎倓埲忋偑梫媮偝傟傞丅偙偺傛偆側崅慄検偲側傞偲丄壥幚偦嵷偺傛偆側悈暘娷検偺懡偄傕偺偱偼徠幩偵傛偭偰惗偢傞梀棧婎偺塭嬁偑戝偒偔丄徠幩廘偺敪惗丄怓慺傗價僞儈儞偺暘夝偑挊偟偔婲傞偺偱栤戣偑懡偄丅偄傑壥廯偺嶦嬠偺堦椺偲偟偰昅幰傜乮侾俋俇係乯偑僀僠僑僕儏乕僗偵徠幩偟偨応崌偺惉暘曄壔傪傒傞偲恾俆乕侾俁偺傛偆偵側傞丅偡側傢偪侽丏俆乣俀俵俼偱僀僠僑偺傾儞僩僔傾儞怓慺偼媫懍偵暘夝偝傟丄娨尦宆傾僗僐儖價儞巁偼巁壔宆偵堏峴偡傞丅傑偨怸梩傜乮侾俋俇侾乯偼僫僣儈僇儞僕儏乕僗偵侾丏俆俵倰倕倫傪徠幩偡傞偲丄桳婡巁丄摐丄傾儈僲巁偼曄壔偟側偄偑丄僼儔僶僲儞偍傛傃傾僗僐儖價儞巁偼尭彮偡傞偲偟偰偄傞丅
丂偲傕偐偔崅慄検偺徠幩偱偼傢傟傢傟偵偲偭偰岲傑偟偔側偄暃斀墳偑婲傞偺偱丄偙偺杊巭曽朄偲偟偰丄梀棧婎偺庴梕懱偺揧壛丄扙巁慺丄徠幩忲棷丄壛擬嶌梡丄掅壏壓徠幩丄扙悈徠幩側偳偑峫偊傜傟偰偄傞丅偙偺偆偪尰嵼偺偲偙傠屻俀幰偑岠壥揑偱偁傞傛偆偱偁傞丅
 |
俙倕倐倕倰倱倧倢倓丆俹丏俠丏丆倕倲丂倎倢丏丗俿倛倕丂倀丏俽丏俙丏俼倕亅
丂倱倕倎倰們倛丂俹倰倧倗倰倎倣丂倧値丂俴倧倵亅俢倧倱倕丂俼倎倓倝倎亅
丂倲倝倧値丂俹倰倧們倕倱倱倝値倗丂倧倖丂俥倧倧倓丆俬値倲丏俰丏俙倫倫亅
丂倢丏俼倎倓丏俬倱倧倲倧倫倕倱丆侾俁丆俁俉俆亅俁俋俁丆乮侾俋俇俀乯
俤倖倖倕們倲倱丂倧倖丂俼倎倓倝倎倲倝倧値丂倧値丂俥倰倳倝倲丂俽倫倧倝亅
丂倢倎倗倕丆俥倧倧倓丂俿倕們倛値倧倢丏侾俉丆侾侾俇侽丆乮侾俋俇係乯丏
俥倧倧倓丂俬倰倰倎倓倝倎倲倝倧値丗俠倧亅俇侽丂俹倢倎値倲丂倝値丂俠倎亅
丂値倎倓倎丆俶倳們倢倕倧値倝們倱丂俀俀丆乮侾侾乯丆俀俉丆乮侾俋俇係乯丏
弿曽朚埨丗壥幚偦嵷偵懳偡傞曻幩慄岠壥丄戞俇夞擔杮傾僀僜僩乕僾夛媍曬暥
丂廤丄俋俁俋乣俋係俁丄乮侾俋俇俆乯丅
攡揷孿巌丗曻幩慄偵傛傞怘昳偺挋憼丄擔杮怘昳岺嬈妛夛戞侾俀夛戝妛僔儞億僕
丂僂儉島墘廤丄侾乣俀丄乮侾俋俇俆乯丅
亙敪夎梷惂亜
俙倱倱倕倢倐倕倰倗倱丆俤丏俙丏俵丏丆倕倲丂倎倢丏丗俤倖倖倕們倲倱丂倧倖
丂俽倧倓倝倳倣丂俵倕倲倎倐倝倱倳倢倫倛倝倲倕丆俙値倲倝倐倝倧倲倝們倱丆丂丂倎値倓丂俧倎倣倣倎丂俬倰倰倎倓倝倎倲倝倧値丂倧値丂倲倛倕
丂俽倛倕倢倖倢倝倖倕丂倧倖丂俹倰倕倫倕倕倢倕倓丂俹倧倲倎倲倧倕倱丂倎倲
丂俼倧倧倣丂俿倕倣倫倕倰倎倲倳倰倕丆丂俙倫倫倢丏俵倝們倰倧倐倝倧倢丏
丂俉丆乮俆乯丆俁侾侾亅俁侾係丆乮侾俋俇侽乯丏
俛倰倧倵値倕倢倢丆俴丏俤丏丆倕倲丂倎倢丏丗俽倲倧倰倎倗倕丂俹倰倧倫亅
丂倕倰倲倝倕倱丂倧倖丂俧倎倣倣倎丂俬倰倰倎倓倝倎倲倕倓丂俹倧倲倎倲倧亅
丂倕倱丆俥倧倧倓丂俿倕們倛倧値倧倢丏侾侾丆俁侽俇亅俁侾俀丆乮侾俋俆俈乯
俢倎倢倢倷値丆俽丏俴丏丆倕倲丂倎倢丏丗俤倶倲倕値倓倝値倗丂俷値倝倧値
丂俽倲倧倰倎倗倕丂俴倝倖倕丂倐倷丂俧倎倣倣倎丂俬倰倰倎倓倝倎倲倝倧値丆
丂俶倳們倢倕倧値倝們倱丂侾俁丆乮係乯丆係俉乣係俋丆乮侾俋俆俆乯丏
丂丂亅亅亅亅亅亅亅亅亅亅亅亅丗俤倖倖倕們倲丂倧倖丂俽倫倰倧倳倲
丂俬値倛倝倐倝倲倝値倗丂俴倕倴倕倢倱丂倧倖丂俧倎倣倣倎丂俬倰倰倎倓倝亅
丂倎倲倝倧値丂倧値丂倲倛倕丂俻倳倎倢倝倲倷丂倧倖丂俷値倝倧値倱丆
丂俹倰倧們丏俙倣倕倰丏俽倧們丏俫倧倰倲丏俽們倝丏俈俁丆俁俋俉亅係侽俇丆
丂乮侾俋俆俋乯丏
俢倳値們倎値丆俢丏俿丏丆倕倲丂倎倢丏丗俽倲倧倰倎倗倕丂俼倧倲丂俽倳倱亅
丂們倕倫倲倝倐倝倢倝倲倷丂倧倖丂俹倧倲倎倲倧丂俿倳倐倕倰倱丂俤倶亅
丂倫倧倱倕倓丂倲倧丂俵倝値倝倣倳倣丂俽倫倰倧倳倲丂俬値倛倝倐倝倲倝値倗
丂俴倕倴倕倢倱丂倧倖丂俬倧値倝倸倝値倗丂俼倎倓倝倎倲倝倧値丆俥倧倧倓
丂俿倕們倛値倧倢丏侾係丆俁俋俆亅係侽侽丆乮侾俋俇侽乯丏
俤倰倰倝値倗倲倧値丆俼丏俥丏丆倕倲丂倎倢丏丗俧倎倣倣倎丂俬倰倰倎倓倝亅丂丂倎倲倝倧値丂倧倖丂俹倧倲倎倧倲倕倱丂倲倧丂俬値倛倝倐倝倲
丂俽倫倰倧倳倲倝値倗丆俠倎値倎倓倝倎値丂俹倧倲倎倲倧丂俬値倓倳倱亅
丂倲倰倷丂俥倝倖倲倛丂俙値値倳倎倢丂俠倧値倖倕倰倕値們倕丆俙倳亅
丂倗倳倱倲丂俈丆侾俋俇侾丏
俫倎値値倎値丆俴丏俽丏丆丗俼倕倱倕倎倰們倛丂倧値丂倲倛倕丂俽們倝亅
丂倕値們倕丂倎値倓丂俿倕們倛値倧倢倧倗倷丂倧倖丂俥倧倧倓丂俹倰倕亅
丂倱倕倰倴倎倲倝倧値丂倐倷丂俬倧値倝倸倝値倗丂俼倎倓倝倎倲倝倧値丆
丂俠倛倕倣丏俹倳倐倢丏俠倧丏丆俬値們丏俶倕倵丂倄倧倰倠丆乮侾俋俆俇乯丏
俬倵倎倲倎丆俿丏丆倕倲丂倎倢丏丗俽倲倳倓倝倕倱丂倝値丂倲倛倕
丂俽倲倧倰倎倗倕丂倧倖丂俠倛倕倱倲値倳倲倱丂倲倰倕倎倲倕倓丂倵倝倲倛
丂俧倎倣倣倎丂俼倎倓倝倎倲倝倧値丆丂俛倳倢倢丏倀値倝倴丏俷倱倎倠倎
丂俹倰倕倖丏俽倕倰丏俛丏俋丆俆俋亅俇俆丆乮侾俋俆俋乯丏
丂亅亅亅亅亅亅亅亅亅亅亅亅亅亅丗俽倲倳倓倝倕倱丂倝値丂倲倛倕
丂俽倲倧倰倎倗倕丂倧倖丂俠倛倕倱倲値倳倲倱丂俿倰倕倎倲倕倓丂倵倝倲倛
丂俧倎倣倣倎丂俼倎倓倝倎倲倝倧値乮俬俬乯丆俛倳倢倢丏俬値倱倲丏
丂俠倛倕倣丏俼倕倱丏俲倷倧倲倧丂倀値倝倴丏俁俋丆侾侾俀亅侾侾俋丆乮侾
丂俋俇侾乯丏
俴倕倵倝倱丆俶丏俥丏丆倕倲丂倎倢丏丗俤倶倲倕値倱倝倧値丂倧倖
丂俽倲倧倰倎倗倕丂俴倝倴倕倱丂倧倖丂俹倧倲倎倲倧倕倱丂倎値倓
丂俷値倝倧値倱丂倐倷丂俠倧倐倎倢倲亅俇侽丂兞亅倰倎倷倱丆俬値倲丏俰丏
丂俙倫倫倢丏俼倎倓丏俬倱倧倲倧倫倕倱丆侾係丆係係俈亅係俆俁丆乮侾俋
丂俇俁乯丏
俵倎倓倱倕値丆俲丏俙丏丆倕倲丂倎倢丏丗俠倕倰倲倎倝値丂俵倧倰倫倛倧亅
丂倢倧倗倝們倎倢丂倎値倓丂俛倝倧們倛倕倣倝們倎倢丂俠倛倎値倗倕倱
丂倝値丂俧倎倣倣倎亅俬倰倰倎倓倝倎倲倕倓丂俠倎倰倰倧倲倱乮俢倎倳們倳倱
丂們倎倰倧倲倎乯丂倎値倓丂俹倧倲倎倲倧倕倱丂乮俽倧倢倎値倳倣
丂倲倳倐倕倰倧倱倳倣丂俴乯丆俼倎倓倝倎倲倝倧値丂俼倕倱倕倎倰們倛
丂侾侽丆係俉亅俇俀丆乮侾俋俆俋乯丏
俵倎倲倛倳倰丆俹丏俛丏丗倁倎倰倝倕倲倷丆丂俢倕倴倕倢倧倫倣倕値倲倎倢
丂俽倲倧倰倎倗倕丂倎値倓丂俢倧倱倕丂俼倎倲倕丂倝値丂俬倰倰倎倓倝倎亅
丂倲倝倧値丂倧倖丂倲倛倕丂俹倧倲倎倲倧丆俶倎倲倳倰倕丂侾俋丆俋俋丆
丂乮侾俋俇俁乯丏
俵倕倲倢倝倱倠倷丆俴丏倁丏丗俛倝倧們倛倕倣倝們倎倢丂俥倳値倓倎倣倕値亅
丂倲倎倢倱丂倖倧倰丂俙倫倫倢倷倝値倗丂俬倧値倝倸倝値倗丂俼倎倓倝倎亅
丂倲倝倧値丂倝値丂俴倕値倗倲倛倕値倝値倗丂倲倛倕丂俹倰倕倱倕倰倴倎亅
丂倲倝倧値丂俹倕倰倝倧倓丂倧倖丂俹倧倲倎倲倧倕倱丆丂倁倕倗倕倲倎亅
丂倐倢倕倱丂倎値倓丂俥倰倳倝倲倱丆丂俙倐倱倲倰丏侾倱倲丏俬値倲丏
丂俠倧値倗倰丏俥倧倧倓丂俽們倝丏丆係俁丆乮侾俋俇俀乯丏
斞捤媊彆傜丗兞慄偵傛傞僞儅僱僊偺朑夎梷惂丄搶嫗搒棫傾僀僜僩乕僾憤崌尋
丂媶強擭曬丂侾丄俈俉乕俉侾丄乮侾俋俇俀乯丏
俵倳倢倢倝値倱丆倂丏俼丏丆倕倲丂倎倢丏丗俿倰倕倎倲倣倕値倲丂倧倖
丂俷値倝倧値倱丂倵倝倲倛丂俧倎倣倣倎丂俼倎倷倱丟俤倖倖倕們倲丂倧倖
丂俢倕倢倎倷丂倐倕倲倵倕倕値丂俫倎倰倴倕倱倲丂倎値倓丂俬倰倰倎倓倝倎亅
丂倲倝倧値丆丂俥倧倧倓丂俿倕們倛値倧倢丏侾俆丆侾俈俉亅侾俈俋丆乮侾俋
丂俇侾乯丏
俵倳倠們倛倝値丆俤丏俶丏丗俛倝倧們倛倕倣倝倱倲倰倷丂倧倖丂俹倧倲倎倲倧
丂俿倳倐倕倰倱丂倓倳倰倝値倗丂俽倲倧倰倎倗倕丏俙倐倎倲倰丏侾倱倲丏
丂俬値倲丏俠倧値倗倰丏俥倧倧倓丂俽們倝丏係俁丆乮侾俋俇俀乯丏
撧椙徣嶰丗兞慄徠幩孖壥摐暘偺挋憼拞偺曄壔丄擾壔俁俆丄係侾俆乕係侾俉丄
丂乮侾俋俇侾乯丅
俶倳倲倲倎倢倢丆倁丏倂丏丆倕倲丂倎倢丏丗俽倧倣倕丂俤倖倖倕們倲倱丂倧倖
丂俧倎倣倣倎丂俼倎倓倝倎倲倝倧値丂倧値丂俽倲倧倰倕倓丂俷値倝倧値倱丆
丂俠倎値倎倓倝倎値丂俰丏俹倢倎値倲丂俽們倝丏係侾丆俉侽俆亅俉侾俁丆
乮侾俋俇侾乯丏
俷倗倎倲倎丆俲丏丆倕倲丂倎倢丏丗俿倛倕丂俤倖倖倕們倲丂倧倖丂俧倎倣倣倎
丂俼倎倓倝倎倲倝倧値倱丂倧値丂俽倫倰倧倳倲丂俹倰倕倱倕倰倴倎倲倝倧値
丂倎値倓丂俬倲倱丂俹倛倷倧倝倧倢倧倗倝們倎倢丂俵倕們倛倎値倝倱倣丂倝値
丂倲倛倕丂俹倧倲倎倲倧丂俿倧倐倕倰丂倎値倓丂倲倛倕丂俷値倝倧値
丂俛倳倢倐丆俛倳倢倢丏俬値倱倲丏俠倛倕倣丏俼倕倱丏俲倷倧倲倧
丂倀値倝倴丏俁俈丆係俀俆亅係俁俇丆乮侾俋俆俋乯丏
俷倞倝倣倎丆俿丏丆倕倲丂倎倢丏丗俧倎倣倣倎丂俬倰倰倎倓倝倎倲倝倧値
丂倧倖丂俷値倝倧値丂俛倳倢倐倱丂倲倧丂俬値倛倝倐倝倲丂俽倫倰倧倳倲亅
丂倝値倗丆丂俙値値倳丏俼倕倫倲丏俼倎倓倝倎倲倝倧値丂俠倕値倲丏俷倱倎亅
丂倠倎丂俹倰倕倖丏係丆侾侾俀亅侾侾係丆乮侾俋俇俁乯丏
俹倕倓倕倰倱倧値丆俽丗俿倛倕丂俤倖倖倕們倲丂倧倖丂俬倧値倝倸倝値倗
丂俼倎倓倝倎倲倝倧値倱丂倧値丂俽倫倰倧倳倲丂俹倰倕倱倕倰倴倎倲倝倧値
丂倎値倓丂俠倛倕倣倝們倎倢丂俠倧倣倫倧倱倝倲倝倧値丂倧倖丂俹倧倲倎亅
丂倲倧倕倱丆丂俥倧倧倓丂俿倕們倛値倧倢丏侾侽丆俆俁俀亅俆俁俇丆
丂乮侾俋俆俇乯丏
俼倳倐倝値丆俛丏俙丏丆倕倲丂倎倢丗俛倝倧們倛倕倣倝們倎倢丂俛倎倱倝倱
丂倧倖丂倲倛倕丂倀倱倕丂倧倖丂兞乕倰倎倷倱丂倲倧丂俤倶倲倕値倓丂倲倛倕
丂俽倲倧倰倎倗倕丂俴倝倖倕丂倧倖丂俹倧倲倎倲倧倕倱丆丂俹倰倧們丏俆亅
丂倲倛丏俬値倲丏俠倧値倗倰丏俛倝倧們倛倕倣丆俵倧倱們倧倵丆俙倳倗丆
丂侾俋俇侾丆倁俬俬俬丆乮侾俇侾亅侾俇俈乯丆俹倂俶亅俹倧倢倝倱倛
丂俽們倝倕値倲倝倖倝們丂俹倳倐倢倝倱倛倕倰丆丂倂倎倰倱倧倵丆
丂乮侾俋俇俁乯丏
俽倎倵倷倕倰丆俼丏俴丏丆倕倲丂倎倢丏丗倁倎倫倧倰倝倸倕倓丂俠倛倕倣倝亅
丂們倎倢丂俬値倛倝倐倝倲倧倰倱丂倎値倓丂俬倰倰倎倓倝倎倲倝倧値丆俿倵倧
丂俶倕倵丂俵倕倲倛倧倓倱丂倧倖丂俽倫倰倧倳倲丂俠倧値倲倰倧倢丂倖倧倰
丂俿倳倐倕倰丂倎値倓丂俛倳倢倐丂俠倰倧倫倱丆俹倰倧丏俙倣倕倰丏俽倧們丏
丂俫倧倰倲丏俽們倝丏俇俈丆俆侾係亅俆俀侾丆乮侾俋俆俇乯丏
俽們倛倰倕倝倐倕倰丆俰丏俽丏丆倕倲丂倎倢丏丗俙丂俽倲倳倓倷丂倧倖
丂倲倛倕丂俛倝倧們倛倕倣倝倱倲倰倷丂倧倖丂俬倰倰倎倓倝倎倲倕倓
丂倫倧倲倎倲倧倕倱丂俽倲倧倰倕倓丂倀値倓倕倰丂俠倧倣倣倕倰們倝倎倢
丂俠倧値倓倝倲倝倧値倱丆俥倧倧倓丂俼倕倱丏俀俁丆係俇係亅係俈俀丆
丂乮侾俋俆俉乯丏
俽倫倎倰倰倧倵丆俙丏俫丏丆倕倲丂倎倢丏丗俬倣倫倰倧倴倕倓丂俽倲倧倰亅
丂倎倗倕丂俻倳倎倢倝倲倷丂倧倖丂俹倧倲倎倲倧丂俿倳倐倕倰倱丂俙倖亅
丂倲倕倰丂俤倶倫倧倱倳倰倕丂倲倧丂俠倧亅俇侽丂俧倎倣倣倎丆俶倳們倢倕亅
丂倧値倝們倱丂侾俀丆乮俉乯丆侾俇亅侾俈丆乮侾俋俆係乯丏
倂倧倰倠倣倎値丆俵丏丆倕倲丂倎倢丏丗俿倛倕丂倀倲倝倢倝倸倎倲倝倧値
丂倧倖丂俬倧値倝倸倝値倗丂俼倎倓倝倎倲倝倧値丂倲倧丂俬値們倰倕倎倱倕
丂倲倛倕丂俽倲倧倰倎倗倕丂俴倝倖倕丂倧倖丂倂倛倝倲倕丂俹倧倲倎倲倧亅
丂倕倱丆丂俥倧倧倓丂俿倕們倛値倧倢丏侾係丆俁俋俆亅係侽侽丆乮侾俋俇侽乯
亙嶦拵嶦棏亜
拑捒榓梇傜丗兞慄徠幩偵傛傞挋憼僋儕奞拵偺嶦尭岠壥丄枹敪昞
亙弉搙挷愡亜
俛倳倰値倱丆俤丏俤丏丆倕倲丂倎倢丏丗俵倎倲倳倰倎倲倝倧値
丂俠倛倎値倗倕倱丂倝値丂俿倧倣倎倲倧丂俥倰倳倝倲倱丂俬値倓倳們倕倓
丂倐倷丂俬倧値倝倸倝値倗丂俼倎倓倝倎倲倝倧値倱丆俥倧倧倓丂俿倕們倛亅
丂値倧倢丏侾侾丆俁侾俁亅俁侾俇丆乮侾俋俆俈乯丏
拑捒榓梇傜丗兞慄徠幩偵傛傞壥幚丒偦嵷偺挋憼偵娭偡傞尋媶乮戞俁曬乯丅墍寍
丂妛夛徍榓俁俉擭搙廐婫戝夛尋媶敪昞丄乮侾俋俇俁乯丅
俥倕倰値倎値倓倕倱丆俽丏俰丏俧丏丆倕倲丂倎倢丏丗俤倖倖倕們倲倱丂倧倖
丂俬倧値倝倸倝値倗丂俼倎倓倝倎倲倝倧値丂倧値丂倲倛倕丂俙們倝倓
丂俵倕倲倎倐倧倢倝倱倣丂倧倖丂俙倫倫倢倕倱乮俠倧倶丂俷倰倎値倗倕
丂俹倝倫倫倝値乯丆俰丏俽們倝丏俥倧倧倓丂俙倗倰倝們丏侾俁丆俀俁亅俀俉丆
丂乮侾俋俇俀乯丏
俫倎値値倎値丆俴丏俽丏丗俼倕倱倕倎倰們倛丂倧値丂倲倛倕丂俽們倝倕値們倕
丂倎値倓丂俿倕們倛値倧倢倧倗倷丂倧倖丂俥倧倧倓丂俹倰倕倱倕倰倴倎亅
丂倲倝倧値丂倐倷丂俬倧値倝倸倝値倗丂俼倎倓倝倎倲倝倧値丆丂俠倛倕倣丏
丂俹倳倐倢丏俠倧丏俬値們丏俶倕倵丂倄倧倰倠丆俶丏倄丏乮侾俋俆俇乯丏
俲倝倲倎倗倎倵倎丆俫丏丆倕倲丂倎倢丏丗俼倕倣倧倴倎倢丂倧倖丂倲倛倕
丂俙倱倲倰倝倗倕値們倷丂倝値丂俲倎倠倝丂倐倷丂俧倎倣倣倎丂俼倎倓倝倎亅
丂倲倝倧値丆俹倰倧們丏俙倕倰丏俽倧們丏俫倧倰倲丏俽們倝丏俉係丆俀侾俁亅
丂俀侾俇丆乮侾俋俇係乯丏
俵倎倱倱倕倷丆俴丏俵丏丆倕倲丂倎倢丏丗俼倎倓倝倎倲倝倧値丂俹倰倧亅
丂們倕倱倱倝値倗丂倧倖丂俥倧倧倓丗俽倧倣倕丂俤倖倖倕們倲倱丂倧倖
丂俧倎倣倣倎丂俼倎倓倝倎倲倝倧値丂倧値丂倲倛倕丂俲倕倕倫倝値倗
丂俻倳倎倢倝倲倷丂倧倖丂俙倫倫倢倕倱丏丆俰丏俙倗倰倝們丏倎値倓
丂俥倧倧倓丂俠倛倕倣丏侾俀丆俀俇俉亅俀俈係丆乮侾俋俇係乯丏
俵倎倶倝倕丆俤丏俠丏丆倕倲丂倎倢丏丗俤倖倖倕們倲倱丂倧倖丂俧倎倣倣倎
丂俼倎倓倝倎倲倝倧値丂倧値丂俼倝倫倕値倝値倗丂倧倖丂俛倎倰倲倢倕倲倲亅
丂倫倕倎倰倱丆俹倢倎値倲丂俹倛倷倱倝倧倢丏俁俋丆俽倳倫倫倢丏倝倶丏
丂乮侾俋俇係乯丏
丂亅亅亅亅亅亅亅亅亅亅亅亅亅亅亅亅丗俽倧倣倕丂俹倛倷倱倝倧倢倧倗倝亅
丂們倎倢丂俤倖倖倕們倲倱丂倧倖丂俧倎倣倣倎丂俬倰倰倎倓倝倎倲倝倧値
丂倧値丂俴倕倣倧値丂俥倰倳倝倲丆俼倎倓倝倎倲倝倧値丂俛倧倲倎値倷丏係丆
丂係侽俆亅係侾侾丆乮侾俋俇係乯丏
弿曽朚埨傜丗兞慄徠幩偵傛傞擾嶻怘昳偺挋憼偵娭偡傞惗棟妛揑尋媶丄戞係夞
丂擔杮傾僀僜僩乕僾夛媍曬暥廤丄侾侽係俆亅侾侽俆侽丄乮侾俋俇俀乯丅
丂亅亅亅亅亅亅亅亅亅亅亅亅亅亅亅亅丗曻幩慄徠幩偵傛傞墍寍怘昳偺挋憼偵
丂娭偡傞尋媶亅僶僫僫偍傛傃僩儅僩偺弉搙偲昳幙偵媦傏偡塭嬁丄擔杮怘昳岺
丂嬈妛夛戞侾侾夞戝夛尋媶敪昞丄乮侾俋俇係乯丅
俼倧倣倎値倝丆俼丏俰丏丗俼倎倓倝倎倲倝倧値丂俹倛倷倱倝倧倢倧倗倷
丂倧倖丂俥倰倳倝倲亅俼倕倱倫倝倰倎倲倝倧値丂倓倳倰倝値倗丂倎値倓
丂俬倣倣倕倓倝倎倲倕倢倷丂俥倧倢倢倧倵倝値倗丂俲倝倢倧倰倎倓丂俢倧倱亅
丂倕倱丂倧倖丂俬倧値倝倸倝値倗丂俼倎倓倝倎倲倝倧値丆俼倎倓倝倎倲倝倧値
丂俛倧倲倎値倷丂係丆俀俋俋亅俁侽俈丆乮侾俋俇係乯丏
俽倎們倳値倠倛倕丆俢丏俲丏丆倕倲丂倎倲丏丗俤倖倖倕們倲倱丂倧倖
丂俧倎倣倣倎丂俼倎倓倝倎倲倝倧値丂俢倧倱倕丆俼倎倲倕丆倎値倓
丂俿倕倣倫倕倰倎倲倳倰倕丂倧値丂倲倛倕丂俿倎倱倲倕丂俹倰倕倖倕倰亅
丂倕値們倕丂倎値倓丂俽倲倧倰倎倗倕丂俴倝倖倕丂倧倖丂俠倕倰倲倎倝値
丂俥倰倳倝倲倱丆倁倕倗倕倲倎倐倢倕倱丆丂倎値倓丂俿倛倕倝倰丂俹倰倧亅
丂倓倳們倲倱丆俹倰倧們丏俙倣倕倰丏俽倧們丏俫倧倰倲丏俽們倝丏丆俈係丆
丂係侾係亅係俀俀丆乮侾俋俆俋乯丏
丂亅亅亅亅亅亅亅亅亅亅亅亅亅亅亅亅亅亅亅丗俹倛倷倱倝倢倧倗倝們倎倢
丂倎値倓丂俠倛倕倣倝們倎倢丂俤倖倖倕們倲倱丂倧倖丂俧倎倣倣倎丂俼倎亅
丂倓倝倎倲倝倧値丂倧値丂俠倕倰倲倎倝値丂俥倰倳倝倲倱丆倁倕倗倕倲倎亅
丂倐倢倕倱丂倎値倓丂俿倛倕倝倰丂俹倰倧倓倳們倲倱丆俹倰倧們丏俙倣倕倰丏
丂俽倧們丏俫倧倰倲丏俽們倝丏俈係丆係俀俁亅係俀俋丆乮侾俋俆俋乯丏
丂亅亅亅亅亅亅亅亅亅亅亅亅亅亅亅亅亅亅亅丗俧倎倣倣倎丂俼倎倓倝倎亅
丂倲倝倧値丂俤倖倖倕們倲倱丂倧値丂俥倰倳倝倲倱丂倎値倓丂倁倕倗倕倲倎亅
丂倐倢倕倱丆俤們倧値丏俛倧倲倎値倷丂侾俆丆俀俉亅俆俇丆乮侾俋俇侾乯丏
憗愳徍傜丗僀僆儞壔曻幩慄徠幩偵傛傞俀丄俁偺擾嶻怘昳偺曐憼偵偮偄偰丄擾嶻
丂媄尋帍丄俉丄係俉乕俆侾丄乮侾俋俇侾乯丅
亙僷僗僣儕僛乕僔儑儞亜
俛倕倰倎倛倎丆俴丏丆倕倲丂倎倢丏丗俧倎倣倣倎丂俼倎倓倝倎倲倝倧値
丂倖倧倰丂俹倧倱倱倝倐倢倕丂俠倧値倲倰倧倢丂倧倖丂俹倧倱倲亅
丂俫倎倰倴倕倱倲丂倧倖丂俙倫倫倢倕倱丆俽倲倰倎倵倐倕倰倰倝倕倱丆
丂俧倰倎倫倕倱丆倎値倓丂俹倕倎們倛倕倱丆俹倛倷倲倧倫倎倲倛倧倢倧倗倷
丂係俈丆係丆乮侾俋俆俈乯丏
丂亅亅亅亅亅亅亅亅亅亅亅亅亅亅亅丗俥倎們倲倧倰倱丂俬値倖倢倳倕値們亅
丂倝値倗丂倲倛倕丂倀倱倕丂倧倖丂俧倎倣倣倎丂俼倎倓倝倎倲倝倧値丂倲倧
丂俠倧値倲倰倧倢丂俢倕們倎倷丂倧倖丂俴倕倣倧値倱丂倎値倓丂俷倰倎値倗亅
丂倕倱丆俹倛倷倲倧倫倎倲倛倧倢倧倗倷丂係俋丆俋侾亅俋俇丆乮侾俋俆俋乯丏
丂亅亅亅亅亅亅亅亅亅亅亅亅亅亅亅丗俤倖倖倕們倲丂倧倖丂俧倎倣倣倎
丂俼倎倓倝倎倝倲倧値丂倧値丂俛倰倧倵値丂俼倧倲丂倎値倓丂俼倛倝倸倧亅
丂倫倳倱丂俼倧倲丂俹倕倎們倛倕倱丂倎値倓丂倲倛倕丂俠倎倳倱倎倢
丂俷倰倗倎値倝倱倣倱丆俹倛倷倲倧倫倎倲倛倧倢倧倗倷丂俆侽丆係俈俆亅
丂係俈俇丆乮侾俋俇侽乯丏
丂亅亅亅亅亅亅亅亅亅亅亅亅亅亅亅丗俬値倖倢倳倕値們倕丂倧倖丂俧倎倣倣倎
丂俼倎倓倝倎倲倝倧値丂俢倧倱倕丂俼倎倲倕丂倧値丂俢倕們倎倷丂倧倖
丂俠倝倲倰倳倱丆俹倕倎倰倱丆俹倕倎們倛倕倱丆倎値倓丂倧値丂俹倕値倝亅
丂們倝倢倢倝倳倣丂倝倲倎倢倝們倳倣丂倎値倓丂俛倧倲倰倷倲倝倱
丂們倝値倕倰倕倎丂倝値丂倴倝倲倰倧丆俹倛倷倲倧倫倎倲倛倧倢倧倗倷
丂俆係丆俈俆俆亅俈俆俋丆乮侾俋俇係乯丏
俠倧倧倫倕倰丆俧丏俵丏丆倕倲丂倎倢丏丗俤倖倖倕們倲丂倧倖丂俧倎倣倣倎
丂俼倎倓倝倎倲倝倧値丆俠倛倕倣倝們倎倢丆倎値倓丂俹倎們倠倎倗倝値倗
丂俿倰倕倎倲倣倕値倲倱丂倧値丂俼倕倖倰倝倗倕倰倎倲倕倓丂俴倝倖倕丂倧倖
丂俽倲倰倎倵倐倕倰倰倝倕倱丂倎値倓丂俽倵倕倕倲丂俠倛倕倰倰倝倕倱丆
丂俥倧倧倓丂俿倕們倛値倧倢丏侾俈丆俉侽侾亅俉侽係丆乮侾俋俇俁乯丏
俢倛倎倢倝倵倎倢丆俙丏俽丏丆倕倲丂倎倢丏丗俬倧値倝倸倝値倗丂俼倎亅
丂倓倝倎倲倝倧値丂倎値倓丂俹倎們倠倎倗倝値倗丂俤倖倖倕們倲倱丂倧値
丂俼倕倱倫倝倰倎倲倧倰倷丂俛倕倛倎倴倝倧倰丂俥倳値倗倎倢丂俧倰倧倵亅
丂倲倛丆丂倎値倓丂俽倲倧倰倎倗倕亅倢倝倖倕丂倧倖丂俹倕倎們倛倕倱丆
丂俹倰倳倣倳倱丂倫倕倰倱倝倕倎丆俼倎倓倝倎倲倝倧値丂俛倧倲倎値倷丂俁丆
丂俈俆亅俉俁丆乮侾俋俇俁乯丏
俰倧倛値倱倧値丆俠丏俥丏丆倕倲丂倎倢丏丗俹倛倷倱倝們倎倢丂倎値倓
丂俽倕値倱倧倰倷丂俿倕倱倲倱丂倧値丂俥倰倕倱倛丂俽倲倰倎倵倐倕倰亅
丂倰倝倕倱丂俽倳倐倞倕們倲倕倓丂倲倧丂俧倎倣倣倎丂俼倎倓倝倎倲倝倧値丆
丂俥倧倧倓丂俿倕們倛値倧倢丏侾俋丆係侾俋亅係俀俁丆乮侾俋俇俆乯丏
斞捤媊彆傜丗俠倧亅俇侽兞慄偵傛傞僀僠僑挋憼帋尡丄搶嫗搒棫傾僀僜僩乕僾
丂憤崌尋媶強擭曬侾丄俉俀乕俉俆丄乮侾俋俇俀乯丅
俵倎倶倝倕丆俤丏俠丏丆倕倲丂倎倢丏丗俤倖倖倕們倲丂倧倖丂俧倎倣倣倎
丂俬倰倰倎倓倝倎倲倝倧値丂倧値丂俿倎倐倢倕丂俧倰倎倫倕倱丆俹倰倧們丏
丂俙倣倕倰丏俽倧們丏俫倧倰倲丏俽們倝丏俉係丆俀俇俁亅俀俇俉丆乮侾俋
丂俇係乯丏
俶倕倢倱倧値丆俲丏俤丏丆倕倲丂倎倢丏丗俽倧倣倕丂俽倲倳倓倝倕倱丂倝値
丂倲倛倕丂倀倱倕丂倧倖丂俬倧値倝倸倝値倗丂俼倎倓倝倎倝倲倧値丂倲倧
丂俠倧値倲倰倧倢丂俛倧倲倧倰倷倲倝倱丂俼倧倲丂倝値丂俿倎倐倢倕
丂俧倰倎倫倕倱丂倎値倓丂俽倲倰倎倵倐倕倰倰倝倕倱丂俹倛倷倲倧倫倎亅
丂倲倛倧倢倧倗倷丂係俋丆係俈俆亅係俉侽丆乮侾俋俆俋乯丏
壀廩傜丗惵壥暔偺曻幩慄挋憼丄搶嫗搒棫傾僀僜僩乕僾憤崌尋媶強擭曬丂俁丄
丂侾俁俈亅侾係係丄乮侾俋俇係乯丅
弿曽朚埨傜丗曻幩慄徠幩偵傛傞墍寍怘昳偺挋憼偵娭偡傞尋媶乕崻嵷椶偺敪夎
丂梷惂暲傃偵壥幚偺捛弉丄挋憼偵懳偡傞曻幩慄岠壥丄擔杮怘昳岺嬈妛夛戞俋
丂夞戝夛敪昞丄乮侾俋俇俀乯丅
俿倰倳倕倢倱倳値丆俿丏俙丏丗俼倎倓倝倎倲倝倧値丂俹倎倱倲倕倳倰倝倸倎亅
丂倲倝倧値丂倧倖丂俥倰倕倱倛丂俥倰倳倝倲倱丂倎値倓丂倁倕倗倕倲倎亅
丂倐倢倕倱丆丂俥倧倧倓丂俿倕們倛値倧倢丏侾俈丆俁俁俇亅俁俁俋丆乮侾俋
丂俇俁乯丏
徠堜棨墱惗傜丗俠倧亅俇侽兞慄徠幩偵傛傞挋憼儕儞僑偺昦奞杊彍丄峅戝擾妛
丂曬侾侾丄俀俁乕俁俀丄乮侾俋俇俆乯丅
丂亅亅亅亅亅丗儕儞僑惵僇價昦嬠偍傛傃僀僠僑奃怓僇價昦嬠偵懳偡傞俠倧亅
丂俇侽兞慄徠幩丄擔杮怉昦妛帍俀俋丄俀俁係乕俀俁俉丄乮侾俋俇係乯丅
俽倎倢倳値倠倛倕丆俢丏俲丏丆倕倲丂倎倢丏丗俹倎們倠倎倗倝値倗
丂俤倖倖倕們倲倱丂倧値丂倲倛倕丂俥倢倎倴倧倳倰丂倎値倓丂俽倛倕倢倖亅
丂倢倝倖倕丂倧倖丂俧倎倣倣倎亅俬倰倰倎倓倝倎倲倕倓丂俥倰倕倱倛
丂俥倰倳倝倲倱丂倎値倓丂倁倕倗倕倲倎倐倢倕倱丆俹倎們倠倎倗倕丂俤値倗丏
丂係丆俁俋亅俆俀丆乮侾俋俆俋乯丏
亙姰慡嶦嬠亜
俵倎倰倎倠倠倝倱丆俹丏丆倕倲丂倎倢丏丗俤倖倖倕們倲倱丂倧倖
丂俠倎倲倛倧倓倕丂俼倎倷丂倎値倓丂俧倎倣倣倎丂俼倎倷丂俬倰倰倎倓倝倎亅
丂倲倝倧値丂倧値丂倲倛倕丂倎値倲倛倧們倷倎値倝値丂俹倝倗倣倕値倲倱
丂倧倖丂俽倲倰倎倵倐倕倰倰倝倕倱丆丂俥倧倧倓丂俼倕倱丏俀係丆俆俀侽亅
丂俆俀俉丆乮侾俋俆俋乯丏
弿曽朚埨丗壥幚偦嵷偵懳偡傞曻幩慄岠壥丄戞俇夞擔杮傾僀僜僩乕僾夛媍曬暥
丂廤丄俋俁俋亅俋係俁丄乮侾俋俇俆乯丅
俷乫倣倕倎倰倎丆俵丏俹丏丗俼倎倓倝倎倲倝倧値丂俠倛倕倣倝倱倲倰倷
丂倎値倓丂俽倲倕倰倝倢倝倸倎倲倝倧値丂倧倖丂俛倝倧倢倧倗倝們倎倢
丂俵倎倲倕倰倝倎倢倱丂倐倷丂俬倧値倝倸倝値倗丂俼倎倓倝倎倲倝倧値倱丆
丂俶倳們倢倕倧値倝們倱丂侾侽丆乮俀乯丆侾俋亅亅俀俁丆乮侾俋俆俀乯丏
怸梩椙擵彆傜丗壞枿姽壥廯偵娭偡傞尋媶乮戞俁曬乯兞慄徠幩偵懳偡傞壔妛惉
丂暘偺曄壔偵偮偄偰丄擾嶻媄尋帍丂俉丄俀俉俆亅俀俉俉丄乮侾俋俇侾乯丅
敀愇惓塸丗傾儊儕僇崌廜崙偵墬偗傞曻幩慄徠幩偵傛傞怘昳偺曐憼尋媶偺尰忬丄
丂擾嶻媄尋帍丂俆丄俈俇乕俉俁丄乮侾俋俆俉乯丅