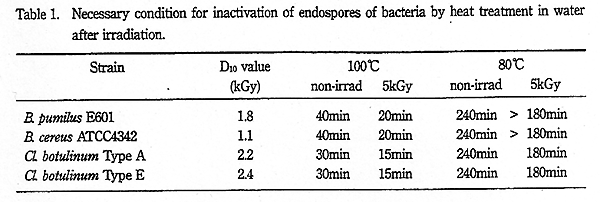
Table 1. Necessary condition for inactivation of endospores of bacteria by heat treatment in water after irradiation.
 |
実験方法
1. 供試菌株
2. 芽胞試料の調整
3. 照射
4. 熱処理
5. 生残菌数の測定
実験結果
1. 乾燥芽胞の放射線感受性
2. 乾燥下でのB. pumilusの耐熱性変化
3. 水共存下での芽胞の耐熱性変化
香辛料等の乾燥食品材料には耐熱性の有芽胞細菌が多量に汚染していることがあり、食品の加工・調理時後に生残し腐敗や食中毒の原因になることがある。乾燥状態の細菌芽胞は著しく耐熱性があり、100℃ では殺菌処理に 10 〜 20 時間必要であり、水存在下でも 30 〜 360 分の処理が必要とされている。一方、食肉製品や魚肉製品の殺菌では中心部の温度を 80℃・20 分以上加熱することが必要とされているが、この加熱条件では細菌芽胞への殺菌効果は全く期待できない。このため、食品衛生法では、これらの食品加工に使用する砂糖、澱粉および香辛料の耐熱性細菌芽胞を 1g 当たり 103 個以下にするように規定している。放射線は香辛料等の乾燥食品材料の殺菌に有効であり、香辛料は 7 〜 10kGy で 1g 当たりの細菌芽胞を 103 個以下に殺菌でき1)、澱粉では 2 〜 5kGy で細菌芽胞をほぼ完全に殺菌できる。しかし、照射後に生残している細菌芽胞の耐熱性についての研究はほとんど行われていない。このため、照射後に生残している細菌芽胞の耐熱性低減の有無を明らかにしておくことは乾燥食品材料の放射線殺菌実用化の上で有意義である。
本研究では、香辛料に多く汚染している Bacillus pumilus と香辛料に比較的多く含まれる B. cereus、土壌などに分布している食中毒性細菌の Clostridium botulinum Type A および Type E を用いて、主に水存在下 100℃ および 80℃ での耐熱性変化について検討した。
供試菌株は B. pumilus E601 株、B. cereus ATCC4342 株、Cl. botulinum Type A 62A 株、Cl. botulinum Type E Biwako 株を用いた。
B. pumilus および B. cereus は芽胞形成用平板培地 (yeast extract 3 g、polypeptone 10 g、glucose 10 g、MnSO4 0.1 g、NaCl 3 g、agar 20 g、水 1 l、pH 7.2) に菌を塗布して 30℃・7 日間培養して芽胞を形成させた。Cl. botulinum Type A および Type E は芽胞形成用液体培地 (tryptone 50 g、polypeptone 5 g、sodium thioglycolate 5 g、glucose 2 g、Na2HPO4 0.35 g、水 1 l、pH 7.0)で 30℃・7 日間培養して芽胞を形成させた。得られた芽胞は 70℃ で 15 分加熱して栄養細胞を不活性化させ、滅菌水で 2 回洗浄した。洗浄した芽胞は 2% ペプトン+ 1% グリセリン水溶液に 108 〜 109 個/ml となるように縣濁し、ガラス繊維濾紙に各 0.2ml 添加し、減圧乾燥した。
ガンマ線源は 10 万キュリー (4.0PBq) の Co-60 板状線源を用い、4.0kGy/h の位置で照射し、照射位置の線量率は Fricke 鉄線量計とアラニン線量計であらかじめ測定しておいた。耐熱性試験用試料はそれぞれ 1、5、10kGy 照射した。
照射した芽胞試料は非照射試料と共に 5ml 蒸留水を入れた試験管、または 10% ペプトン水 5ml を入れた試験管に浸し、シリコセンで栓をした状態で加熱処理をした。乾燥下での耐熱性試験は試験管に芽胞試料を入れたもの、および試験管下部に蒸留水を入れ上部に芽胞試料を添付した状態で加熱処理した。100℃ での加熱処理はシリコンオイル槽で行い、80℃ での加熱処理は恒温水槽で行った。
照射後または加熱処理後の生残菌数は滅菌生理食塩水 (含 0.01% Tween 20) で希釈してから平板培養または MPN 法で測定した。B. pumilus および B. cereus の生残菌数は Difco-nutrient agar 平板培地に出現してくるコロニー数から測定した。Cl. botulinum Type A および Type E の生残菌数は Thioglycolate broth を用い MPN (10本) 法で濁度を示す試験管数から測定した。
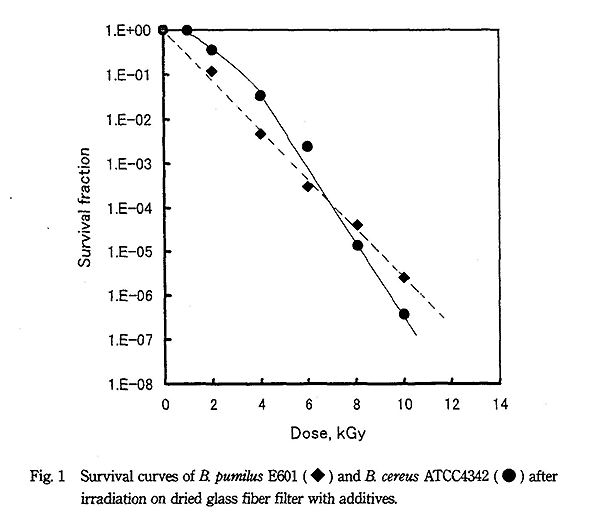
B. pumilus の芽胞を無添加の状態で乾燥すると、ガラス繊維濾紙でもセルロース繊維濾紙でも D10 値は 1.6kGy になる2)。一方、ガラス繊維濾紙にペプトン+グリセリン共存下で乾燥すると D10 値は 1.8kGy になった。B. cereus の芽胞も無添加乾燥での D10 値は 0.9kGy になり、他の Bacillus 属に比べ高い放射線感受性を示した。一方、ペプトン+グリセリン共存下では Fig. 1 に示すように D10 値は大きくなり 1.1kGy になった。
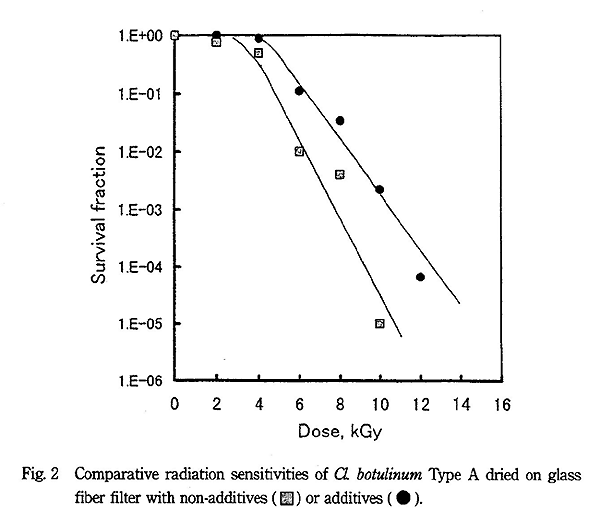
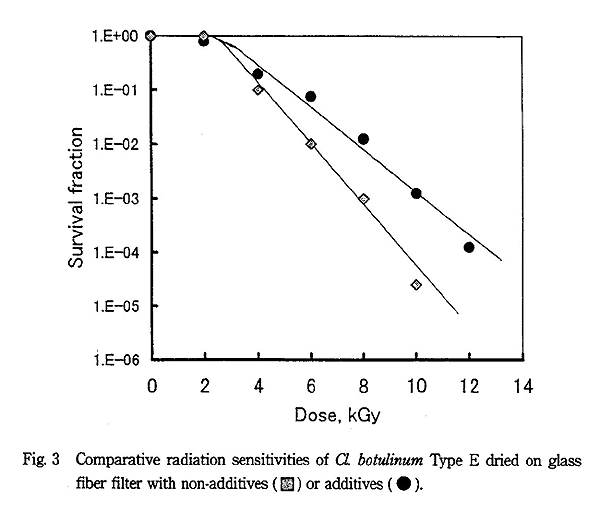
Cl. botulinum は Bacillus 属に比べて放射線耐性が強いことが知られているが、Fig. 2、3 に示すように無添加での D10 値は Type A で 1.5kGy、Type E で 1.7kGy になった。しかし、ペプトン+グリセリン共存下では D10 値が Type A で 2.2kGy、Type E で 2.4kGy と著しく放射線に耐性になった。
各菌種の生存曲線は B. pumilus が指数関数型を示すのに対し、他の菌種は全てシグモイド型を示し、Cl. botulinum の両株は 5kGy でもほとんど殺菌効果を示さないほど大きな肩を有していた。
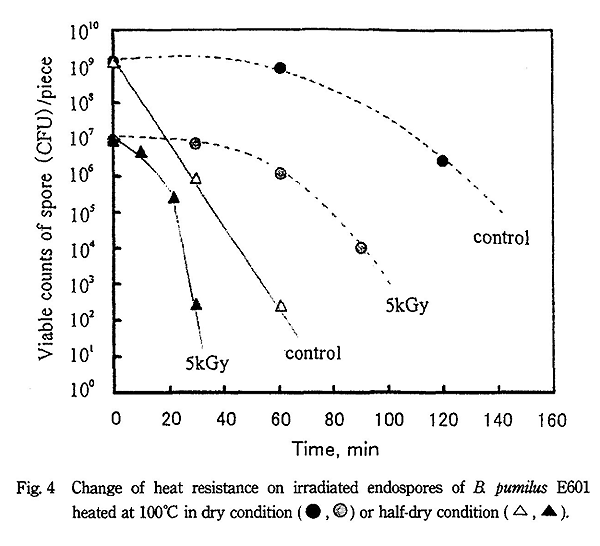
Fig. 4 に示すように、B. pumilus の芽胞を乾燥下で加熱すると 100℃・2 時間処理後でも生残菌数はほとんど低減せず、5kGy 照射試料でも加熱による殺菌促進効果はほとんど認められなかった。一方、高湿度の半乾燥下で加熱すると、非照射芽胞でも約 1 時間処理で生残菌数は著しく減少した。また、5kGy 照射すると約 30 分で著しく菌数が低減し殺菌促進効果が認められた。しかし、乾燥下では長時間加熱しないと殺菌効果が得られないため、他の菌種についての耐熱性は検討しなかった。
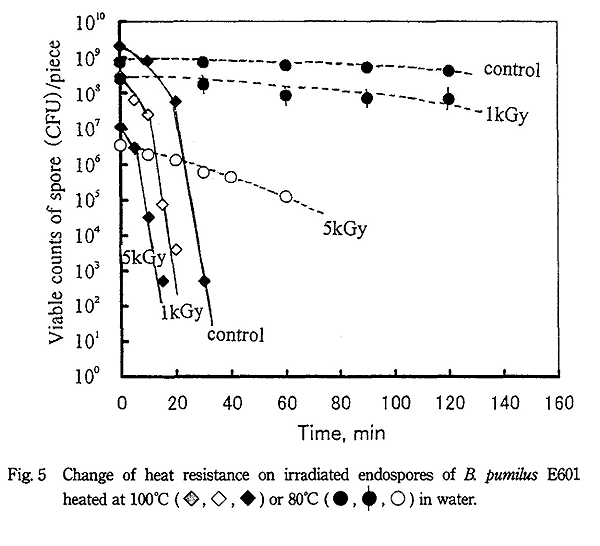
B. pumilus 芽胞の試料片を蒸留水または 10% ペプトン水に浸けてから 100℃ で加熱処理すると、Fig. 5 に示すように非照射芽胞は約 40 分で殺菌され、蒸留水とペプトン水による差は認められなかった。一方、1kGy 照射芽胞は 30 分で殺菌され、5kGy 照射芽胞は 20 分で殺菌され、加熱による殺菌促進効果が認められた。また、80℃ の加熱処理では非照射芽胞と 1kGy 照射芽胞は 2 時間処理でもほとんど殺菌されないのに対し、5kGy では殺菌促進効果が若干認められた。B. cereus の場合も非照射芽胞は 100℃ では約 40 分で殺菌されたのに対し、5kGy では 20 分で殺菌された。また、80℃ での照射による耐熱性変化も B. pumilus と同様の傾向を示した。
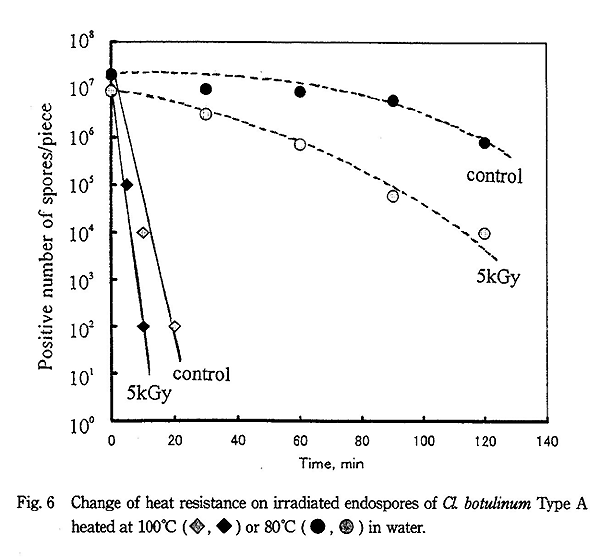
Cl. botulinum Type A の場合、Fig. 6 に示すように非照射芽胞の耐熱性は B. pumilus や B. cereus より低く 100℃ では約 30 分で殺菌された。一方、5kGy 照射された芽胞は放射線での殺菌効果はほとんどなかったが、100℃ の加熱処理では約 15 分で殺菌され殺菌促進効果が認められた。しかし、80℃ での加熱処理では 5kGy 照射または 10kGy 照射でも殺菌促進効果はほとんど認められなかった。Cl. botulinum Type E の場合も Type A とほぼ同じ傾向を示した。
これらの結果をまとめると Table 1 に示すようになり、各菌種の芽胞とも水共存下・100℃ の加熱処理では照射による殺菌促進効果が著しく認められたが、80℃ では殺菌促進効果はほとんど認められなかった。
本研究では、各菌株とも 5kGy 照射した芽胞は水存在下での加熱による殺菌促進効果が 100℃ で認められた。放射線処理後に生残している芽胞が水存在下・100℃ での耐熱性が低減する理由は、放射線で受けた DNA 損傷が加熱によりさらに拡大されるためと考えられる。しかし、80℃ では DNA 損傷の拡大効果はほとんどないと思われる。一方、あらかじめ細菌芽胞を 70℃ で前処理しても放射線感受性促進効果は認められないが、80℃ 以上では放射線感受性が促進されると報告されている3)。このことは、放射線と加熱による殺菌促進効果が DNA 鎖切断と熱変成の相乗効果によることを示しているように思われる。
ところで、食品の加工・調理において耐熱性細菌の汚染は食品衛生の上で大きな問題である。細菌芽胞の場合、水存在下・100℃ で 40 分以上加熱処理しないと殺菌が困難であり、この条件では食品の栄養成分の破壊や食味低下が問題になる。また、80℃・20 分での殺菌処理では加工食品に汚染している細菌芽胞ばかりでなく、一般細菌の殺菌も不十分である。
放射線による乾燥食品材料の殺菌の場合、細菌芽胞の殺菌線量は 2 〜 10kGy 必要である。放射線で殺菌処理された乾燥食品材料を用いた加工食品が加熱調理時にさらに殺菌促進効果が認められれば加熱時間の短縮、食品衛生の向上につながることが期待できる。
本研究の結果では 100℃ 以上の加熱により殺菌促進効果が明確に認められ、必要加熱時間が 5kGy で半分以下に短縮することができた。しかし、80℃ では照射による殺菌促進効果はわずかしか認められないため、乾燥食品材料中の細菌芽胞数を十分低減してから食品加工に使用するべきであろう。なお、80℃ 以下の温度では照射芽胞ばかりでなく一般細菌の殺菌促進効果も全く期待できないであろう。一方、強力な食中毒菌である Cl. botulinum Type A や Type E は芽胞形成細菌の中で強い放射線耐性を示したが、熱処理との組み合わせによる殺菌促進効果は他の芽胞形成細菌より顕著なことが明らかになった。このことは、好気性の芽胞形成細菌の計測により得られた殺菌線量が嫌気性の芽胞形成細菌にも適用できることを示していると思われる。
1) Muhamad L. J., H. Ito, H. Watanabe and N.Tamura : Agric. Biol. Chem., 50(2), 347-355 (1986).
2) 伊藤 均 : 医科器械学, 60(10), 409-475 (1990).
3) 山崎邦郎, 岡 充, 伊藤宣夫, 飯塚義助 : 食品照射, 1(1), 27-33 (1966).
(2001 年 5 月 18 日受理)
|
 |
 |
 |
 |
 |
 |
|
|