戞侾乮倎乯恾 抝偟傖偔偺俠倛倢倧倰倧倫倛倷倢倢倎倱倕丂倎們倲倝倴倝倲倷
 |
丂曻幩慄丄偲偔偵倃慄徠幩偵傛偭偰丄攏楅彃偺敪夎傪梷惂偟丄挋憼婜娫偺墑挿傪偼偐傞帋傒偼丄偡偱偵奺崙偵偍偄偰幚梡壔偺抜奒偵擖偭偰偄傞丅敪夎梷惂偺栚揑偵揔偭偨幚梡慄検乮俈乣侾俆倠倰倎倓乯偱偼丄塰梴壙偦偺懠晽枴側偳偵塭嬁偑傒傜傟偢丄傑偨怘昳岺嬈尨椏偲偟偰偺昳幙側偳偵傕丄壗摍埆塭嬁偑擣傔傜傟偰偄側偄丅偝傜偵丄廳梫側栤戣偲偟偰偺挿婜摦暔帞堢偵傛傞埨慡惈偺妋擣帋尡偵偍偄偰傕丄桳奞嶌梡偑偁傞偲偄偆曬崘偼尒摉偨傜側偄丅
丂偟偐偟丄堦曽丄奺崙偵偍偄偰傕丄偄傑偩偵徠幩怘昳偱偁傞偙偲傪幆暿偡傞曽朄偼尒弌偝傟偰偄側偄丅傑偨丄尰忬偱偼丄婛偵埨慡惈偑妋擣偝傟偰偄傞慄検丄偡側傢偪丄揔惓慄検偱徠幩偑巤偝傟偰偄傞偐斲偐傪幆暿偡傞曽朄偲偟偰偼丄徠幩帪偵儔僪僇儔乕傪揬晅偡傞曽朄埲奜偵偼丄
丂攏楅彃偼丄惗慛擾嶌暔偱偁傝丄偦偺昳庬丄嶌晅娐嫬丄廂妌帪婜丄挋憼婜娫偁傞偄偼忦審偑晄柧偺応崌偼丄帋椏嵦庢偺曽朄傗懳徠暔偺嵦傝曽偵傛傞曄摦偺曽偑丄偼傞偐偵戝偒偄応崌傕峫偊傜傟傞偺偱丄暔幙儗儀儖傪栤戣偲偡傞側傜偽丄偁傞惉暘偺徚挿偁傞偄偼丄偁傞惉暘偺怴偟偄惗惉偑傒傜傟偨帪偵丄偼偠傔偰徠幩偺桳柍傪妋擣偡傞偙偲偑偱偒傞偺偱偼側偄偐偲峫偊傜傟傞丅
丂埲壓丄傢傟傢傟偑峴偭偰偒偨徠幩張棟偺専抦朄偵娭偡傞婎慴揑尋媶偲偦偺寢壥偵偮偄偰曬崘偡傞丅
丂廂妌帪婜丗徍榓係係擭俇寧俀俇乣俀俉擔
丂嶻丂丂抧丗挿嶈導
丂徠幩帪婜丗徍榓係係擭俈寧俉擔
丂挋憼忦審丗壏搙丟俀侽乣俀俀亷
丂丂丂丂丂丂幖搙丟俆侽乣俉侽亾
丂丂丂丂丂丂婜娫丟徍榓係係擭俈寧侾侽擔乣徍榓係係擭侾俀寧侾俀擔
丂廂妌帪婜丗徍榓係係擭俋寧侾俆乣俀侽擔
丂嶻丂丂抧丗杒奀摴
丂徠幩帪婜丗徍榓係係擭侾侽寧俀俉擔
丂挋憼忦審丗壏搙丟俀侽乣俀俀亷
丂丂丂丂丂丂幖搙丟俆侽乣俉侽亾
丂丂丂丂丂丂婜娫丟徍榓係係擭侾侾寧係擔乣徍榓係俆擭俁寧俀侾擔
丂廂妌帪婜丗徍榓係俁擭俋寧俀俆擔
丂嶻丂丂抧丗杒奀摴
丂徠幩帪婜丗徍榓係俁擭侾侾寧侾俁乣侾係擔
丂挋憼忦審丗壏搙丟乮幒壏乯
丂丂丂丂丂丂幖搙丟惉傝備偒
丂丂丂丂丂丂婜娫丟徍榓係俁擭侾侾寧俀俆擔乣徍榓係俆擭俆寧侾侽擔
丂廂妌帪婜丗徍榓係俁擭俋寧俀俆擔
丂嶻丂丂抧丗杒奀摴
丂徠幩帪婜丗徍榓係俁擭侾侾寧侾俁乣侾係擔
丂挋憼忦審丗壏搙丟幒壏
丂丂丂丂丂丂幖搙丟惉傝備偒
丂丂丂丂丂丂婜娫丟徍榓係俁擭侾侾寧俀俆擔乣徍榓係俆擭俆寧侾侽擔
丂擔杮尨巕椡尋媶強
丂丂崅嶈尋媶強丂丂丂丂丂兞慄乮俠倧亅俇侽乯
丂壗傟傕俈丄侾俆偍傛傃俁侽倠倰倎倓偺徠幩
丂夠宻偺捀晹偍傛傃掙晹偱塰梴惉暘偺暘晍偑堎側傞偙偲偐傜廲偺椫愗傝乮拞怱幉偵偦偭偰乯傪俁屄懱偐傜暯嬒偵嵦庢偟偨丅偨偩偟丄俹倕倰倧倶倝倓倎倱倕偵偮偄偰偼墶偺椫愗傝乮拞怱幉偵捈妏偵乯偲偟偨丅
丂倂倧倛倢倗倕倣倳倲倛朄仏侾乯偵廬偄丄倫俫俆丏侽偺俙們倕倲倎倲倕丂倐倳倖倖倕倰拞偱峺慺塼侾倣倢偑俁俈亷俁侽暘偱暘夝偟偆傞侾亾壜梟惈揵暡塼検傪媮傔偨乮俢俁俈並俁侽乫偱昞帵乯丅傑偨丄倣倕倱倲倕們倠倕倷仏侾侽乯偵傛傞倱倲倎倰們倛丂倎倗倎倰丂倫倢倎倲倕傪梡偄偰丄娙曋朄偵偮偄偰傕専摙偟偨丅
丂俛倕倱倱倕倷亅俴倧倵倰倷朄仏俀乯偵傛傝丄巁惈偍傛傃傾儖僇儕惈壓偱丄倫亅値倝倲倰倧倫倛倕値倷倢丂倫倛倧倱倫倛倎倲倕傪婎幙偲偟偨偲偒偵惗惉偡傞倫亅値倝倲倰倧倫倛倕値倧倢傪係侾侽倣兪偱斾怓掕検偟偨丅
丂俼倕倝倲倣倎値亅俥倰倎値倠倕倢朄仏俁乯偵婎偯偄偰丄倎倢倎値倝値倕傑偨偼丄倎倫倎倰倎倲倝們丂倎們倝倓丂偲丂俀亅倠倕倲倧倗倢倳倲倎倰倝們丂倎們倝倓丂傪婎幙偲偟偰丄惗惉偡傞俹倷倰倳倴倝們丂倎們倝倓傑偨偼丄倧倶倎倢倧倎們倕倲倝們丂倎們倝倓丂傪倓倝値倝倲倰倧倫倛倕値倷倢丂倛倷倓倰倎倸倝値倕偵傛傝敪怓丄斾怓掕検偟偨丅
丂俫倧倢倓倕値仏係乯偺曽朄偵廬偄丄帋椏傪俆侽亾埲忋偺傾僙僩儞擹搙偱丄扙悈暡枛偲側偟丄偙傟傪傾僙僩儞俆侽亾丄僋僄儞巁娚徴塼乮侽丏係俵乯丄俠倳亅們倛倢倧倰倧倫倛倷倢倢乮俀倣倗乛倣倢乯偺婎幙拞乮倫俫俉丏侽乯偵寽戺偟丄俀俆亷偱俀丄俆丄俉丄俀侽帪娫偺僀儞僉儏儀乕僩屻丄暘夝惗惉暔偺們倛倢倧倰倧倫倛倷倢倢倝倓倕傪暘暿偟丄斾怓掕検偟偨丅
丂俿倷倰倧倱倝値倎倱倕仏侾俁乯乮俠倰倕倱倧倢倎倱倕偲俠倎倲倕丂們倛倧倢倎倱倕乯丄俹倕倰倧倶倝倓倎倱倕仏侾俁乯側偳偵偮偄偰傕専摙偟偨丅
丂悈梟惈憤摐検偼丄彍僞儞僷僋屻丄倎値倲倛倰倧値倕朄偱應掕偟偨丅
丂俽倧倣倧倗倷倝亅俶倕倢倱倧値偺曄朄仏俆乯偵傛傝掕検偟丄僌儖僐乕僗偲偟偰昞帵偟偨丅
丂乮倝乯偐傜乮倝倝乯傪嵎堷偒丄潋摐偲偟偰昞帵偟偨丅
丂倂丏倂倧倰倎倴倓倕倠倎倰仏俇乯傜偺曽朄偵廬偄丄夁儓僂慺巁巁壔偵傛傝惗惉偟偨丂倣倎倢倧値亖倓倝倎倢倓倕倛倷倓倕丂偲丂倲倛倝倱倐倎倰倐倝倲倳倰倝們丂倎們倝倓偲偺弅崌暔傪斾怓掕検偟偨丅
丂俼倧倕偺俀丏係亅倓倝値倝倲倰倧倫倛倕値倷倢丂倛倷倓倰倎倸倝値倕丂朄偵俿俴俠傪暪梡丄夵椙偟偨曽朄仏俈丄俉乯傪梡偄偰掕検偟偨丅側偍丄廬棃偺俼倧倕偺尨朄偱偼巁壔宆傾僗僐儖價儞巁偲偟偰掕検偟偨傕偺偑丄俿俴俠偱偼恀偺傾僗僐儖價儞巁偲偟偰偼嵀愓掱搙偟偐懚嵼偟側偄偙偲偑柧偐偲側偭偨偺偱丄憤傾僗僐儖價儞巁偲偟偰掕検丄昞帵偟偨丅
丂傾儈僲巁偺倲倷倰倧倱倝値倕偼俠倕倰倝倧倲倲倝仏俋乯傜偺曽朄丄僋儘儘僼傿儖丄儕僌僯儞條暔幙乮僼儘儘僌儖僔儞棸巁朄乯丄僕儀儗儕儞巁仏侾侾乯丄柍婡儕儞仏侾俀乯丄傾儞儌僯傾懺拏慺丄傾儖僨僸僪椶偼忢朄偵傛傝専摙偟偨丅
丂帋椏俁丄係偺挋憼屻俁僇寧栚偺敪夎婜偺寢壥偼丄戞侾昞偺捠傝偱偁傞丅旕徠幩丄俈倠倰倎倓徠幩偺傕偺偼丄侾俆倠倰倎倓丆俁侽倠倰倎倓徠幩偟偨傕偺傛傝傕丄傗傗妶惈偑崅偄偑丄挋憼弶婜偵偼丄帋椏侾丄俀傪梡偄偨応崌偱傕丄壗傟傕妶惈偑掅偔丄嵎堎偼擣傔傜傟側偐偭偨丅傑偨丄僕儀儗儕儞梟塼乮侽丏侽俆俵乯偵俀係帪娫怹捫偟偰傕丄帋椏拞偺峺慺晩妶壔偼擣傔傜傟偢丄俙倣倷倢倎倱倕偺妶惈傪斾妑偡傞偙偲偼丄桳岠側庤抜偲偼峫偊傜傟側偄丅
| 丂丂丂 昳丂庬 |
丂丂丂丂丂丂丂 丂慄丂丂丂検丂 |
俢丂俁俈並 丂丂俁侽乫 |
| 丂丂丂 丂抝丂 丂偟丂 丂傖丂 丂偔丂 丂係丂 丂丂丂 |
丂侽乮旕徠幩乯 |
丂侽丏侾丂 |
| 丂俈丂倠倰倎倓 |
丂侽丏侾丂 |
|
| 侾俆丂丂乂丂丂 |
丂侽丏侾丂 |
|
| 俁侽丂丂乂丂丂 |
丂侽丏侽俈 |
|
| 丂丂丂 丂擾丂 丂椦丂 丂侾丂 丂崋丂 丂俁丂 丂丂丂 |
丂侽乮旕徠幩乯 |
丂侽丏侽俈 |
| 丂俈丂倠倰倎倓 |
丂侽丏侽俈 |
|
| 侾俆丂丂乂丂丂 |
丂侽丏侽俆 |
|
| 俁侽丂丂乂丂丂 |
丂侽丏侽俆 |
丂俙們倝倓丂倫倛倧倱倫倛倎倲倎倱倕丂偺應掕寢壥偼丄戞俀昞偵帵偟偨丅傑偨丄屄懱娫偺峺慺妶惈抣偺乽偽傜偮偒乿傪傒傞偨傔丄抝偟傖偔傪慖傃丄俆屄懱偺悢抣傪媮傔偨偲偙傠丄奺屄懱偺妶惈抣偵傒傜傟傞昗弨岆嵎偼栺俈亾偱偁傝丄俆亾偺婋尟棪傪傒傞偲偒怣棅抣偼暯嬒亇侾俈丏俇亾偺暆偲側偭偨丅廬偭偰戞俀昞偺寢壥偐傜丄徠幩偺塭嬁傪丄偙傟偵傛偭偰悇應偡傞偙偲偼崲擄偱偁傞偲巚傢傟傞丅
|
丂亂俛倕倱倱倕倷亅俴倧倵倰倷丂倳値倝倲丂俵乛倛倰乛倗乮倖倰倕倱倛丂倲倝倱倱倳倕乯亃 |
| 丂丂丂丂 昳庬丂丂 丂丂丂丂 |
丂慄丂丂検丂 丂丂丂丂丂丂 乮倠倰倎倓乯 |
丂丂丂徠幩屻偺挋憼婜娫丂丂丂 |
||
| 侽丂儢寧 |
侾丂儢寧 |
俀丂儢寧 |
||
| 丂丂丂丂 丂丂丂丂 搰丂丂尨 丂丂丂丂 丂侾丂丂 丂丂丂丂 丂丂丂丂 |
丂丂侽丂丂丂 丂丂丂丂丂丂 丂丂俈丂丂丂 丂丂丂丂丂丂 丂侾俆丂丂丂 丂丂丂丂丂丂 丂俁侽丂丂丂 |
侽丏俈俋 丂丂丂丂 侽丏俆俈 丂丂丂丂 侽丏俇俉 丂丂丂丂 侽丏俋侽 |
侽丏俇俉 丂丂丂丂 侽丏俈俋 丂丂丂丂 侽丏俈俋 丂丂丂丂 侽丏俇侽 |
侽丏俇俉 丂丂丂丂 侽丏俆俈 丂丂丂丂 侽丏俇俉 丂丂丂丂 侽丏俇俉 |
| 丂丂丂丂 丂丂丂丂 擾椦侾崋 丂丂丂丂 丂俁丂丂 丂丂丂丂 丂丂丂丂 |
丂丂侽丂丂丂 丂丂丂丂丂丂 丂丂俈丂丂丂 丂丂丂丂丂丂 丂侾俆丂丂丂 丂丂丂丂丂丂 丂俁侽丂丂丂 |
丂亅丂丂 丂丂丂丂 丂亅丂丂 丂丂丂丂 丂亅丂丂 丂丂丂丂 丂亅丂丂 |
丂亅丂丂 丂丂丂丂 丂亅丂丂 丂丂丂丂 丂亅丂丂 丂丂丂丂 丂亅丂丂 |
侽丏俁俁 丂丂丂丂 侽丏俁侾 丂丂丂丂 侽丏俁侾 丂丂丂丂 侽丏俀俋 |
| 丂丂丂丂 丂丂丂丂 抝偟傖偔 丂丂丂丂 丂係丂丂 丂丂丂丂 丂丂丂丂 |
丂丂侽丂丂丂 丂丂丂丂丂丂 丂丂俈丂丂丂 丂丂丂丂丂丂 丂侾俆丂丂丂 丂丂丂丂丂丂 丂俁侽丂丂丂 |
丂亅丂丂 丂丂丂丂 丂亅丂丂 丂丂丂丂 丂亅丂丂 丂丂丂丂 丂亅丂丂 |
丂亅丂丂 丂丂丂丂 丂亅丂丂 丂丂丂丂 丂亅丂丂 丂丂丂丂 丂亅丂丂 |
侽丏俀係 丂丂丂丂 侽丏俀侽 丂丂丂丂 侽丏俀俁 丂丂丂丂 侽丏俀俋 |
丂搰尨傪慖傃丄倲倰倎値倱倎倣倝値倎倱倕偺戙昞揑側峺慺妶惈偲偟偰俧俷俿丏俧俹俿傪應掕偟丄戞俁昞偵偦偺寢壥傪帵偟偨丅徠幩捈屻偼丄徠幩張棟傪巤偟偨傕偺偺曽偑崅偄抣傪帵偟偨丅偟偐偟丄杮應掕偺傛偆偵丄奺孮偐傜弌棃傞偩偗戝偒偝偺懙偭偨屄懱傪俆屄慖傫偱應掕偟偰傕丄奺屄懱娫偺偽傜偮偒偼丄戝懱亇俈亾乮暯嬒抣乯偱偁偭偨丅偟偨偑偭偰丄俆亾偺婋尟棪傪傒傞偲偒丄怣棅抣偺暆偼亇侾俈亾偲側傝丄枹抦偺帋椏偺峺慺妶惈偑崅偔偲傕丄偙傟偑徠幩張棟傪巤偟偨傕偺偱偁傞偐斲偐傪悇掕偡傞偙偲偼偱偒側偄丅傑偨丄擾椦侾崋偍傛傃抝偟傖偔偵偮偄偰傕忋弎摨條側孹岦傪帵偟偨丅
|
丂乮扨埵丗俲倎倰倣倕値倱丂倳値倝倲倱丄俵乛倣倢乛倗乯 |
| 丂慄丂丂検丂 丂丂丂丂丂丂 乮倠倰倎倓乯 |
丂丂丂俧丂俷丂俿丂丂丂 |
丂丂丂俧丂俹丂俿丂丂丂 |
||||
| 侽儢寧 |
侾儢寧 |
俁儢寧 |
侽儢寧 |
侾儢寧 |
俁儢寧 |
|
| 丂丂丂侽丂丂 丂丂丂丂丂丂 丂丂丂俈丂丂 丂丂丂丂丂丂 丂丂侾俆丂丂 丂丂丂丂丂丂 丂丂俁侽丂丂 |
丂俇俆 丂丂丂 丂俇俆 丂丂丂 丂俈俉 丂丂丂 丂俋俆 |
丂俋俆 丂丂丂 丂俋侽 丂丂丂 丂俈俉 丂丂丂 侾侽侽 |
丂俋俆 丂丂丂 丂俋俆 丂丂丂 丂俈俉 丂丂丂 丂俋侽 |
丂係俁 丂丂丂 丂係俇 丂丂丂 丂俇侽 丂丂丂 丂俈侽 |
丂俇俆 丂丂丂 丂俇俆 丂丂丂 丂俇侽 丂丂丂 丂俈俆 |
丂俇俆 丂丂丂 丂俈侽 丂丂丂 丂俇侽 丂丂丂 丂俇侽 |
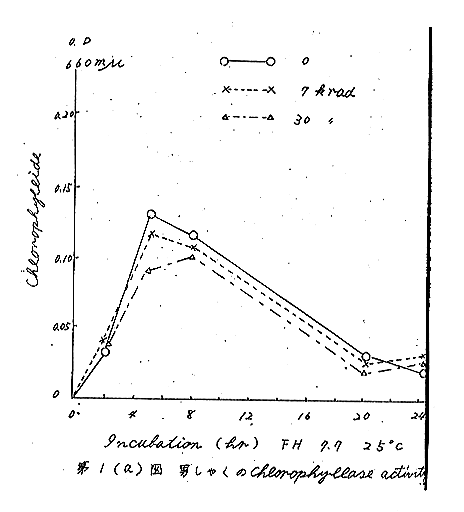
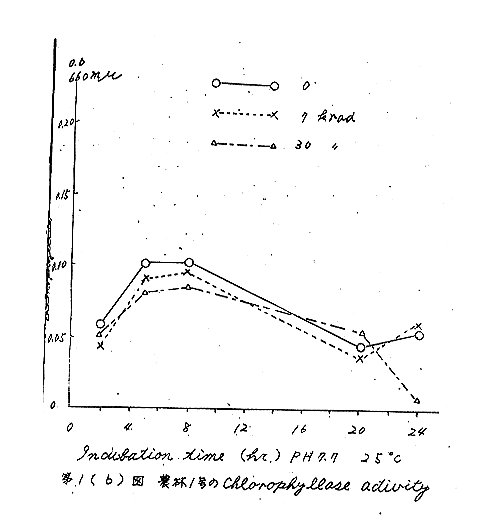
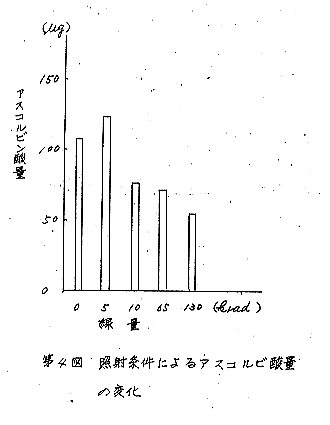
丂峺慺妶惈偑嵟傕崅偄敪夎婜乮挋憼屻俁僇寧乯偺寢壥傪戞侾乮倎乯丄乮倐乯恾偵帵偟偨丅恾偺傛偆偵僀儞僉儏儀乕僩偺弶婜偵偼丄徠幩偟偨傕偺偺曽偑掅偄孹岦傪帵偟偨偑丄偙傟偼検揑偵岆嵎抣撪偵擖傞傕偺偱偁傞丅
丂傑偨丄傾僙僩儞暡枛偺戙傢傝偵倖倰倕倱倛丂倲倝倱倱倳倕乮僋僄儞巁僜乕僟梟塼拞偱儂儌僕僫僀僘偟偨傕偺乯傪梡偄偨応崌傕摨條偱偁傝丄僕儀儗儕儞梟塼乮俆倫倫倣乯俀係帪娫怹捫偟偨傕偺偱傕曄壔偼傒傜傟側偐偭偨丅偙偺峺慺偼丄慹惢偱丄偐偮丄傾僙僩儞梟塼拞偱偺傒妶惈傪帵偡偑丄偦偺斀墳偼抶偔丄偟偐傕惛惢偡傞偵廬偭偰丄妶惈傪幐偆偺偱丄専抦朄偲偟偰偼揔摉側曽朄偲偼巚傢傟側偄丅
|
 |
丂俹倕倰倧倶倝倓倎倱倕偼俫俀俷俀乮俀亊侾侽丒俤乮亅俁乯俵乯丄俙倱們倧倰倐倝們丂倎們倝倓乮侾侽丒俤乮亅俇乯俵乯丄俠倎倲倕們倛倧倢倎倱倕丂偼俠倎倲倕們倛倧倢乮俆亊侾侽丒俤乮亅俉乯俵乯丆丄倎倱們倧倰倐倝們丂倎們倝倓乮俀丏俆亊侾侽丒俤乮亅俈乯俵乯俠倰倕倱倧倢倎倱倕丂偼俹亅們倰倕倱倧倢乮侾亊侾侽丒俤乮亅係乯俵乯丄俙倱們倧倰倐倝們丂倎們倝倓乮侾亊侾侽丒俤乮亅係乯俵乯傪偦傟偧傟婎幙偲偟偰丄攏楅彃乮抝偟傖偔乯偺旂晹偍傛傃擏幙晹偵暘偗偰俛倕値倸倝倓倝値倕怓慺朄偱應掕偟偨丅俹倕倰倧倶倝倓倕妶惈偼旂晹偵傒傜傟徠幩屻侾僇寧乮俁侽倠倰倎倓乯偱旕徠幩偺傕偺偺栺俀攞偵払偟丄俀偍傛傃俁僇寧屻偱傕傗傗崅偄孹岦傪帵偟偨偑丄擏幙晹偱偼妶惈偼嬌傔偰掅偔丄徠幩丄旕徠幩偺娫偱敾慠偲偟偨嵎堎偼擣傔傜傟側偐偭偨丅堦曽丄俠倎倲倕們倛倧倢倎倱倕偍傛傃們倰倕倱倧倢倎倱倕偼壗傟傕旂晹丄擏晹偲傕偵徠幩丄旕徠幩娫偵桳堄嵎偼擣傔傜傟側偐偭偨丅
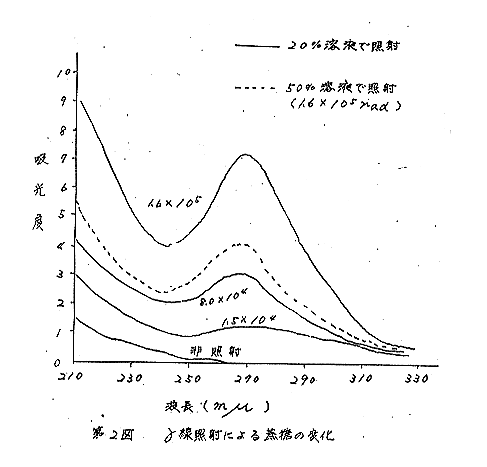
丂潋摐乮俆亾丄俀侽亾乯梟塼偵丄兞慄傪侾丏俆亊侾侽丒俤乮係乯丄俉丏侽亊侾侽丒俤乮係乯丄侾丏俇亊侾侽丒俤乮俆乯倰倎倓丆俀俉亷偱徠幩偟偨婎慴幚尡偺寢壥傪戞俀恾偵帵偟偨丅
丂偙傟偼俀俇俆乣俀俈侽倣兪偺娫偺僌儖僐乕僗丄僼儔僋僩乕僗側偳偺扨摐椶偲堦抳偡傞僺乕僋偑丄侾丏俆亊侾侽丒俤乮係乯倰倎倓乮侾俆倠倰倎倓乯偱尰傟丄偙偺僺乕僋偼徠幩慄検傪憹偡偵廬偭偰憹戝偡傞偙偲傪帵偟偰偄傞丅傑偨丄擹搙偑婬敄側梟塼偱丄倰倎倓倝倧倢倷倱倝倱偑憹嫮偝傟傞偐偳偆偐妋偐傔傞偨傔偵丄俆亾媦傃俀侽亾梟塼偱偺徠幩寢壥傪斾妑偟偨偑丄偙偺擹搙偱偼丄俀侽亾梟塼偺倰倎倓倝倧倢倷倱倝倱偺曽偑戝偒偐偭偨丅
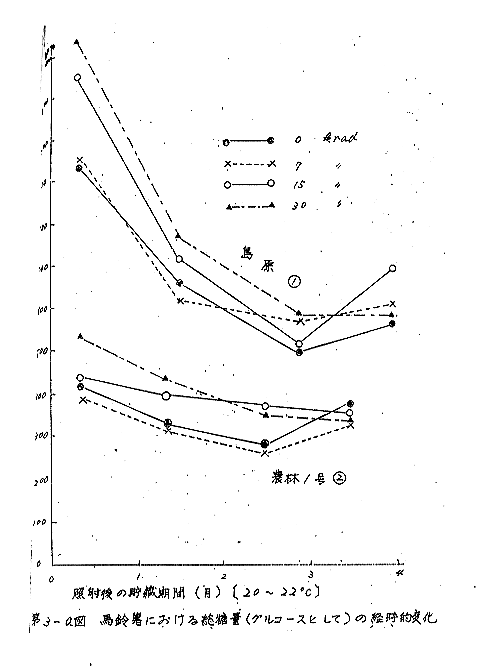
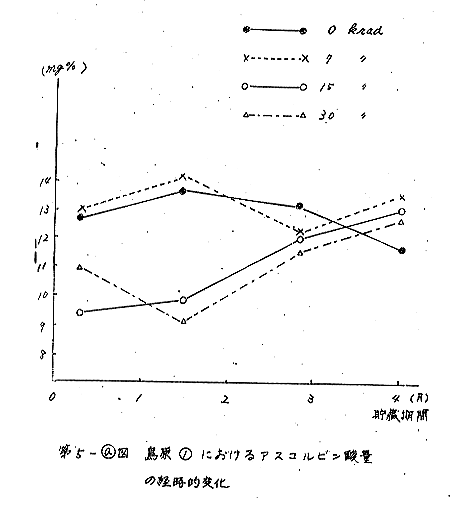
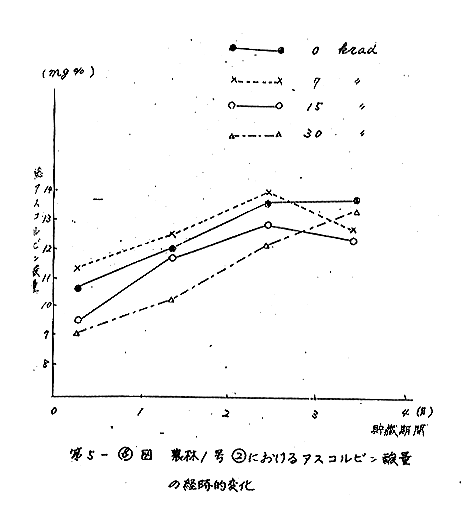
丂偙傟傜偺寢壥偐傜丄徠幩偵傛傝娨尦摐検偑憹壛偡傞傕偺偲悇掕偟丄徠幩屻敪夎乮旕徠幩孮乯傑偨偼晠攕乮徠幩孮乯偑偼偠傑傞傑偱偺婜娫偺昳庬暿偵傛傞娨尦摐偍傛傃旕娨尦摐検偺曄壔傪戞俁乮倎乯丆乮倐乯丆乮們乯恾偵帵偟偨丅憤摐検乮戞俁乮倎乯恾乯偼丄徠幩屻丄俁僇寧埲撪偱偼憹壛偺孹岦偑傒傜傟偨偑丄偙傟偼丄偲偔偵娨尦摐偺憹壛偵傛傞傕偺偲偼巚傢傟側偐偭偨乮戞俁乮倐乯恾乯丅
丂挋憼婜娫拞偺摐娷検曄壔偼擾椦侾崋乮俀丄俁乯偍傛傃抝偟傖偔乮係乯偼戝懱摨偠傛偆側孹岦傪帵偟丄帪婜偵傛傞曄摦偑彫偝偄偺偵懳偟偰丄搰尨乮侾乯偱偼娨尦摐媦傃旕娨尦摐偲傕偵擾椦侾崋偺栺俀攞検傪娷桳偟丄帪婜偵傛傞曄摦偼挊偟偐偭偨丅偟偐偟丄挋憼屻俀僇寧傪宱傞偲徠幩慄検娫偵堦掕偺孹岦偼擣傔傜傟側偔側偭偨丅偲偔偵昳庬傗挋憼婜娫偺晄柧側傕偺偵偁偭偰偼丄摐偺娷桳検偺懡彮偐傜徠幩偺桳柍傪悇掕偡傞偙偲偼晄壜擻偱偁傞偲巚傢傟傞丅
丂傑偨丄倓倕倧倶倷丂倱倳倗倎倰偵偮偄偰偼丄潋摐偍傛傃僌儖僐乕僗偺俀侽亾悈梟塼偵侾俆丄俉侽丄侾俇侽倠倰倎倓徠幩偟偨傕偺偍傛傃丄徠幩攏楅彃乮抝偟傖偔擾椦侾崋亅俈丄侾俆丄俁侽倠倰倎倓徠幩乯偦偺傕偺丄偍傛傃丄俁侽倠倰倎倓徠幩攏楅彃乮抝偟傖偔乯偐傜挷惢偟偨揵暡拞偐傜偼丄偄偢傟傕専弌偱偒側偐偭偨丅
 |
 |
 |
 |
丂婎慴揑側幚尡偲偟偰丄傾僗僐儖價儞巁俆亾儊僞椨巁梟塼乮侾丏侽俈兞乛倣倢乯傪丄偁傜偐偠傔僲乕儕僢僩偱巁壔偟偨傕偺傪彫僇僢僾偵俀倣倢偢偮偲傝丄師偺忦審偱丄俠倧亅俇侽偵傛偭偰徠幩偟偨丅奺慄検暿偵傛傞帋椏偼丄俢俶俹俫朄偱掕検偟丄偐偮俿俴俠忋偺僗億僢僩偺埵抲傪妋擣偟偨丅
丂徠幩偵傛傞傾僗僐儖價儞巁偺曄壔偼戞係恾偵帵偟偨丅
丂傑偨丄俿俴俠忋偵偼傾僗僐儖價儞巁偍傛傃倓倕倛倷倓倰倧倎倱們倧倰倐倝們丂倎們倝倓丂偲堦抳偡傞僗億僢僩埲奜偺傕偺偼専弌偝傟側偐偭偨丅
丂側偍丄幚嵺偺忦審偲偼堎偵偟偰丄梊傔巁壔傪峴偭偨偺偼丄幚尡偺娙堈壔偺偨傔偱偁傝丄傑偨丄僲乕儕僢僩偵傛傞巁壔偼姰慡偵巁壔偝傟傞傕偺偱偼側偄偑丄廘慺悈傗偦偺懠偵傛傞巁壔曽朄偲斾妑偡傞偲偒丄偙偺曽朄偑嵟傕埨掕偟偨寢壥傪摼偨丅
丂偟偐偟丄戞係恾偵傒傜傟傞傛偆偵丄旕徠幩偺傕偺傛傝俆倠倰倎倓徠幩偟偨傕偺偺曽偑崅偄悢抣傪帵偡偺偼丄僲乕儕僢僩偱巁壔偝傟側偐偭偨傾僗僐儖價儞巁偑徠幩偵傛傝巁壔偟偨傕偺偲巚傢傟傞丅偙偺恾偐傜侾侽倠倰倎倓偱傾僗僐儖價儞巁偵曄壔偑偁傜傢傟傞偑丄侾俁侽倠倰倎倓偺徠幩偱傕丄栺俆侽亾偑巆懚偡傞偲偙傠偐傜丄傾僗僐儖價儞巁帺懱偼徠幩偵傛偭偰丄偦傟掱晄埨掕偺傕偺偱偼側偄傛偆偱偁傞丅
丂師偵搰尨侾丄擾椦侾崋俀偵偮偄偰丄徠幩屻偺挋憼婜娫偵傒傜傟傞傾僗僐儖價儞巁偺曄壔傪宱帪揑偵應掕偟偨乮戞俆恾乯丅椉昳庬偲傕徠幩屻丄挋憼婜娫偺弶婜偵偼旕徠幩偵斾傋丄傾僗僐儖價儞巁娷検偼丄掅壓偺孹岦傪帵偡偑丄俁僇寧埲屻偵偼偙偺孹岦偼傒傜傟側偔側偭偨丅偟偐偟丄俈倠倰倎倓掱搙偺徠幩偱偼旕徠幩偺傕偺偲敾暿偡傞偙偲偼弌棃側偐偭偨丅
丂傑偨丄抝偟傖偔係偱傕丄徠幩屻侾僇寧偱旕徠幩帋椏偑丄侾俁丏俈倣倗亾傪帵偡偺偵懳偟丄俁侽倠倰倎倓帋椏偱偼俇丏俇倣倗亾偺掅偄抣偱偁偭偨丅偟偐偟丄俁僇寧屻偵偼丄旕徠幩帋椏偼敪夎偟偼偠傔丄傾僗僐儖價儞巁検偼丄侾侾丏俀倣倗亾傪帵偟偨偺偵懳偟丄俁侽倠倰倎倓帋椏偼侾俆丏侽倣倗亾偲曄傢傝丄忋婰俀昳庬偲摨條側宱夁傪帵偟偨丅
| 丂丂丂丂慄丂丂丂丂丂検丂丂丂丂 |
丂嫍丂丂丂棧丂 |
丂帪丂丂娫丂 |
| 俆丂丂亊侾侽丒俤乮俁乯丂倰倎倓 丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂 侾丂丂亊侾侽丒俤乮係乯丂倰倎倓 丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂 俇丏俆亊侾侽丒俤乮係乯丂倰倎倓 丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂 侾丏俁亊侾侽丒俤乮俆乯丂倰倎倓 |
侾侽丂丂丂們倣 丂丂丂丂丂丂丂 丂俆丏俇丂們倣 丂丂丂丂丂丂丂 丂俀丂丂丂們倣 丂丂丂丂丂丂丂 丂俀丂丂丂們倣 |
丂侾侽丂暘丂 丂丂丂丂丂丂 丂丂俇丂暘丂 丂丂丂丂丂丂 丂丂俆丂暘丂 丂丂丂丂丂丂 丂侾侽丂暘丂 |
 |
 |
 |
丂抝偟傖偔帋椏偵偮偄偰傾儖僨僸僪椶丄傾儞儌僯傾懺拏慺丄柍婡椨丄倲倷倰倧倱倝値倕丆倗倝倐倐倕倰倕倢倢倝們丂倎們倝倓傪暘愅偟偨丅傑偨丄寀岝摂壓偱俀廡娫曻抲偟偰們倛倢倧倰倧倫倛倷倢倢偺惗惉傪専偟偨丅
丂傾儖僨僸僪椶丄傾儞儌僯傾懺拏慺偼徠幩丄旕徠幩偺娫偱慡偔嵎堎偼擣傔傜傟側偐偭偨丅傑偨丄柍婡椨丄倲倷倰倧倱倝値倕偼慄検暿娫偵戝偒側嵎堎偼擣傔傜傟側偐偭偨乮戞俆昞乯丅
丂俧倝倐倐倕倰倕倢倢倝們丂倎們倝倓偵偮偄偰偼丄壗傟偺帋椏偐傜傕専弌偱偒側偐偭偨丅偟偐偟丄倗倝倐倐倕倰倕倢倢倝們丂倎們倝倓偺昗昳偲偼俿俴俠忋偱堦抳偟側偄偑丄寀岝傪桳偡傞僗億僢僩傪専弌偟偨丅偙偺僗億僢僩偼俁侽倠倰倎倓徠幩偺傕偺偺曽偑旕徠幩偺傕偺偵斾傋偰寀岝偑嫮偐偭偨偑丄寀岝媧廂嬋慄偺僷僞乕儞偵偼丄嵎堎偑擣傔傜傟側偐偭偨丅
丂師偵丄攏楅彃傪捠忢偺寀岝摂壓偵敇偡偲偒丄們倛倢倧倰倧倫倛倷倢倢丂傪惗惉偡傞丅偙偺椢旂尰徾乮俧倰倕倕値倝値倗乯偼丄兞慄徠幩偵傛傝梷惂偝傟傞偲偄偆曬崘偑偁傞偑丄徠幩屻侾儢寧偺帋椏偱偼旕徠幩偍傛傃俈倠倰倎倓徠幩帋椏傪侾侽侽偲偡傞偲偒丄侾俆偍傛傃俁侽倠倰倎倓偺帋椏偼俆俆丏俁偱偁偭偨丅偟偐偟丄挋憼擔悢偺宱夁偲偲傕偵椢旂尰徾偼晄妶敪偲側傞丅傑偨丄僕儀儖儕儞張棟丄扽巁僫僩儕僂儉張棟傪巤偟偰傕椢旂尰徾偼懀恑偝傟側偐偭偨丅
| 丂慄丂丂丂丂丂検丂 |
倫倝丂乮倣倗亾乯 |
倲倷倰倧倱倝値倕丂乮倣倗亾乯丂 |
| 旕丂徠丂幩丄丂丂夎 丂丂丂丂丂丂丂丂丂 旕丂徠丂幩丄丂夠宻 丂丂丂丂丂丂丂丂丂 丂俈倠倰倎倓丂夠宻 丂丂丂丂丂丂丂丂丂 侾俆倠倰倎倓丂夠宻 丂丂丂丂丂丂丂丂丂 俁侽倠倰倎倓丂夠宻 |
丂丂俁侽丂丂丂丂 丂丂丂丂丂丂丂丂 丂丂侾係丂丂丂丂 丂丂丂丂丂丂丂丂 丂丂侾俇丂丂丂丂 丂丂丂丂丂丂丂丂 丂丂侾俋丂丂丂丂 丂丂丂丂丂丂丂丂 丂丂俀侽丂丂丂丂 |
丂丂丂丂丂丂俀俉丂丂丂丂丂丂丂 丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂 丂丂丂丂丂丂俆俀丂丂丂丂丂丂丂 丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂 丂丂丂丂丂丂俆俀丂丂丂丂丂丂丂 丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂 丂丂丂丂丂丂俆侾丂丂丂丂丂丂丂 丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂 丂丂丂丂丂丂俈侽丂丂丂丂丂丂丂 |
丂偙傟傑偱偺徠幩攏楅彃偵偍偗傞尋媶寢壥偐傜丄兞慄張棟偵傛傝柧傜偐偵曄壔偡傞傛偆側峺慺妶惈傪應掕偡傞曽朄偼丄奺屄懱暿偵傛傝僶儔僣僉偑戝偒偄偲偄偆揰偐傜傕丄徠幩丄旕徠幩偺専抦朄偲偟偰偼揔惈傪寚偔傛偆偵巚傢傟傞丅
丂堦曽丄塰梴惉暘偺曄壔偵偮偄偰偼丄徠幩屻丄挋憼婜娫偑俁儢寧傑偱偼丄旕徠幩偺傕偺偲偁傞掱搙偺悢抣揑側嵎堎偼擣傔傜傟傞偑丄偙傟傜偼怴偟偄惉暘偺惗惉傗丄偁傞惉暘偺徚幐偲偄偭偨傕偺偱偼側偔丄検揑側憹尭偵偡偓側偄偨傔丄偙偺悢抣偑旕徠幩偲徠幩張棟偟偨傕偺偲偺娫偱嬌抂側嵎堎偑側偗傟偽専抦朄偲偟偰偺梡傪側偝側偄丅傑偨丄挋憼婜娫偑俁儢寧傪夁偓傞偲丄偦偺嵎堎偼擣傔傜傟側偔側傞偺偱丄偙偺庬偺暔幙儗儀儖偱偺専抦朄偲偟偰偼丄尰抜奒偱偼晄揔摉偱偁傞丅偟偨偑偭偰丄偙傟偐傜偺栤戣偲偟偰丄兞慄徠幩偵傛傞敪夎梷惂偑懠敪揑媥柊傪梌偊傞倣倎倢倕倝們丂倛倷倓倰倎倸倝倓倕丂倲倕倲倰倎們倛倢倧倰倧値倝倲倰倧倐倕値倸倕値倕側偳偺壔妛栻嵻偵傛傞敪夎梷惂偲嫟捠偟偨婡峔偵傛傞傕偺偐斲偐丄傑偨丄嵎堎偑偁傞傕偺偱偁傞側傜偽丄偄偐側傞揰偵偁傞偺偐傪夝柧偡傞偙偲偑昁梫偱偁傞丅
丂傑偨丄兞慄徠幩偵傛傞敪夎梷惂婡嶌偑丄媥柊婡峔偺墑挿偱偼側偔丄扨偵敪夎嵶朎偺攋夡偵傛傞傕偺偲悇掕偡傞側傜偽慻怐妛揑偵斾妑偡傞専抦朄偑傛傝岠壥揑偱偁傞偲峫偊傜傟傞丅偨偲偊偽丄宍懺妛揑専嶕偼栜榑偱偁傞偑丄慻怐愗曅忋偱偺壔妛揑偁傞偄偼峺慺妛揑斀墳傪棙梡偡傞偙偲偵傛偭偰丄杮曬偺寢壥傪偝傜偵徻嵶偵夝柧偟偆傞傕偺偲巚傢傟傞丅偟偐偟忋弎偟偨捠傝丄慻怐妛揑専抦朄傕傑偨丄壔妛栻嵻張棟傪巤偟偨傕偺偲斾妑専摙偡傞偙偲偑昁梫偲側傞傕偺偲峫偊傜傟傞丅
丂侾乯丂俰丏倂倧倛倢倗倕倣倳倲倛丏俛倝倧們倛倕倣丏倅丏俋丆侾乮侾俋侽俉
丂丂丂丂丂乯
丂俀乯丂俷丏俙丏俛倕倱倱倕倷丂倕倲丂倎倢丏丆俰丏俛倝倧倢丏俠倛倕倣丏丆
丂丂丂丂侾俇係丆俁俀侾乮侾俋係俇乯
丂俁乯丂俽丏俼倕倝倲倣倎値丂倕倲丂倎倢丏丆俙倣丏俰丏俠倢倝値丏俹倎倲倛
丂丂丂丂丏丆俀俉丆俆俇乮侾俋俆俈乯
丂係乯丂俵丏俫倧倢倓倕値丏俛倝倧們倛倕倣丏俰丏丆俈俉丆俁俆俋乮侾俋俇侾
丂丂丂丂乯
丂俆乯丂俵丏俽倧倣倧倗倷倝丆俰丏俛倝倧倢丏俠倛倕倣丏丆侾俋俆丆侾俋乮侾
丂丂丂丂俋俆俀乯
丂俇乯丂倂丆俽丏倂倎倰倎倴倓倕倠倎倰丂倕倲丂倎倢丏丆俰丏俛倝倧倢丏俠倛
丂丂丂丂倕倣丏丆俀俁係丆侾俋係俆乮侾俋俆俋乯
丂俈乯丂俫丏俫倎倱倱倕倢倯倝倱倲丂倕倲丂倎倲丏丆俙們倲倎丂俠倛倕倣丏俽
丂丂丂丂們倎値倓丏侾俈丆俆俀俋乮侾俋俇俁乯
丂俉乯丂俽倲倰倧倛倕們倠倕倰丏丆倅丏俴倕倐倕値倱倣倝倲倲倕倢丏俥倧倰倱
丂丂丂丂們倛丏丆侾侾俉丆俁俋係乮侾俋俇俁乯
丂俋乯丂俧丏俠倕倰倝倧倲倲倝丂倕倲丂倎倢丏丆俛倝倧們倛丏丆俇俇丆俇侽俈
丂丂丂丂乮侾俋俆俈乯
侾侽乯丂俰丏俵倕倱倲倕們倠倷丏丂俙値倎倢丏俛倝倧倢丏丆俁侽丆侾俋侽乮侾
丂丂丂丂俋俇俋乯
侾侾乯丂俰丏俧倧倰倓倧値丂倕倲丂倎倢丏丆丂俰丏俙倗倰丏俥倧倧倓丂俠倛倕
丂丂丂丂倣丏丆侾俇丆俆俀侽乮侾俋俇俉乯
侾俀乯丂拞懞摴摽丄擾壔丄俀係丄侾乮侾俋俆侽乯
侾俁乯丂俼丏俹丏俥丏俧倰倕倗倧倰倷丏丆俛倝倧們倛倕倣丂俰丏丆侾侽侾丆俆
丂丂丂丂俉俀乮侾俋俇俇乯
丂丂丂丂乮媨栘崅柧丄枔栰宨揟丄搚揷夒巕丄塹摗挬巕丄拞懞抦巕乯
|
|