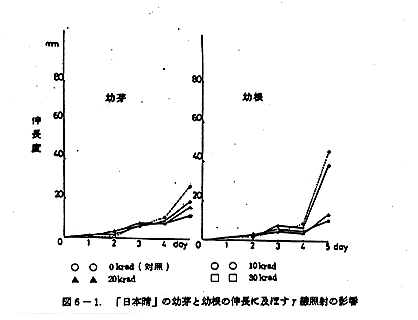
図6-1 「日本晴」の幼芽と幼根の伸長に及ぼすγ線照射の影響
 |
食品衛生上の見地から照射処理の有無ならびにその適正照射量の有無を検知、鑑別する方法の開発研究が要請されるにいたった。昭和47年現在までのところ定められた低線量条件で照射された穀物に関しては、国の内外を問わず、適切な検知方法に関する報告は見当たらないのが実情である。しかしながら、限定された条件下で、示唆に富む実験結果を得たので報告する。
米
a)こしひかり-昭和45年秋収穫
-昭和46年6月28日照射
b)日 本 晴-昭和46年秋収穫
-昭和46年12月2日照射
原研で照射処理後、予研に送付された。研究室においては、20℃恒温器内に保蔵したが、湿度管理はされていない。
日本原子力研究所・高崎研究所のγ線照射装置(Co-60)を用い、10、20、30krad照射を行った。
対照非照射試料及び各照射試料について経時的にその変化を検討した。
ベルトラン法、レイン・エイノン法およびソモギー法による検討に加え、薄層クロマトグラフィー[担体 シリカゲルG,溶媒酢酸エチル:イソプロパノール:水(65:23:12)で展開後、アニスアルデヒドー硫酸で発色]を採用した。
本来還元性を示さない澱粉が、照射によってグリコシド結合が切れて何等かの変化を生ずる可能性を予想し、照射試料について水溶性糖分除去処理後、経時的に還元力を測定した。
粉砕試料10gに40℃純水100mlを加え振盪後、40℃低温保持し、30分及び60分後に振盪したのち濾過して濾液を得た。濾液50mlにフェノールフタレイン指示薬を加え、N/50水酸化ナトリウム標準液で滴定し、照射処理米の澱粉が示す酸度(アルカリ度)を検討した。
粉砕試料10gをとり、そのベンゼン抽出液にエタノールを加え、フェノールフタレインを指示薬として0.178N水酸化カリウム標準液で滴定し、照射試料の脂肪酸度を比較検討した。
粉砕照射試料からソックスレー法によって抽出した脂質区分について、常法によりケン化価及びウィイス法によりヨウ素価を測定し、比較検討した。
供試種子は、予め2%次亜塩素酸ナトリウム中に室温で1時間浸漬して表面殺菌した。50粒づつペトリ皿上に蒔種し、30℃恒温器中に保持して、毎日発芽した粒を分別し、3日間の発芽率で比較した。
発芽率測定と同様に処理して発芽させ、5日間にわたって芽と主根の長さを測定し、それぞれ1粒あたりの平均値で記録した。
前記(4)と同様に発芽させ、2日目の主根伸長先端から5mmのところで切断したものを試料として、次の方法によって組織標本を作製した。
(イ) 酢酸カーミン染色による押しつぶし法:
試料をエタノール:酢酸(3:1)で固定した。IN塩酸で60℃、10分間処理したのち、酢酸カーミンで染色し、スライドガラス上で押しつぶした。
(ロ) ハイデンハイン氏ヘマトキシリン染色によるパラフィン切片法:
試料をナワシン液(1%クロム酸:氷酢酸:ホルマリン 10:1:4)で固定し、パラフィン包埋したのちに、5~7μの厚さに切片を作り、ハイデンハイン氏ヘマトキシリンで染色した。
上記2つの方法を併用し、顕微鏡下で細胞分裂後期のブリッジ(染色体橋)の有無を測定した。
また、パラフィン切片を用いて、根端の中心柱がまっすぐ縦に入った縦断面を選び、根冠を除いた140μmの間にある部位の細胞について、染色体異常の有無を観察した。
a) 発芽時のホスファターゼ活性変化、トランスアミラーゼ活性変
化、及びアミラーゼ活性について測定。
b) ヘミセルローズの変化(酸、アルカリ、ジアスターゼによる水
解)観察
c) 炊飯のレオロジー
結果として、実用照射線量条件下における米の照射において、昭和45年前後での技術水準をもって照射の有無もしくは照射線量の強弱を迅速簡易に検出し得る程の変化は生じないことが明らかになった。
すなわち、照射米穀の還元糖、非還元糖及びデオキシ糖、澱粉の酸度(アルカリ度)及び還元力、脂質区分の脂肪酸度、ケン化価及びヨウ素価、ヘミセルローズの変化(酸アルカリあるいはジアスターゼによる水解度)、炊飯のレオロジー的観察などにあっては、照射量の如何を問わず非照射区との間に有意の差は認められなかった。発芽時におけるホスファターゼ、トランスアミナーゼ、アミラーゼ等の酵素活性は、発芽率の上昇につれて高まるように見えるが、この変化は照射量の多少によるというよりは、供試穀物の品種、収穫後の貯蔵条件、照射までの経過時間などに大きく影響を受けることが判った。
このように化学的あるいは生化学試験によっては明確に照射の有無を判別することは困難であった。
そこで、発芽に及ぼす照射の影響について検討した結果を次に記す。
発芽率について:蒔種3日目における発芽率を表6-1に示す。
全般的に、今回の実験におけるような低線量照射条件にあっては、線量の高低による影響はほとんど認められない。「日本晴」と「こしひかり」とでは、後者の発芽率が著しく劣るようにみえるが、これは品種の差というよりは、実験条件の差というべきと考えられる。すなわち、見掛上発芽率の劣る「こしひかり」は収穫後8~9ヶ月後しかも梅雨期を過ぎて照射したものであるが、収穫後適切な時期における照射が重要なことを示唆する結果を得た。
| 品 種 |
線 量 (krad) |
|||
| 0 |
10 |
20 |
30 |
|
| 日 本 晴 |
100 |
100 |
98 |
98 |
| こしひかり |
72 |
32 |
36 |
42 |
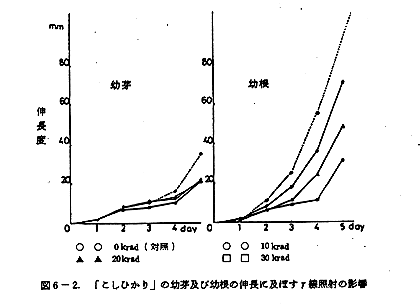
蒔種後5日目にいたる発芽伸長の状況を図6-1,及び図6-2に示した。
発芽及びその後の幼芽の伸長に対して10~30krad低線量照射の範囲において、対照とした非照射群との間に照射後4日目までは差は認められなかった。5日目にやや照射群に幼芽伸長阻害が見られた。その傾向は品種の差によらないようである。
発根及びその後の幼根の伸長に対する照射の影響を見ると、「日本晴」と「こしひかり」では大きく異なる。「こしひかり」では幼芽の場合と同様に4日目まで対照群と照射群に差はみられず、5日目になって多少照射(20~30krad)の影響と考えられる幼根の伸長阻害が見られた。「日本晴」の場合は、照射2日目から照射線量の高低に応じた幼根伸長阻害が観察された。しかし、全般的に幼根の伸長は、良好であり、30krad照射群ですら「こしひかり」の対照群と大差ない伸長度を示した。両品種のこの大差は、「日本晴」の方が「こしひかり」に比較して鮮度の高いうちに照射処理を受け、かつ発育試験を実施したことによるのかも知れない。この点を確認するためには、再試験を要する。
米の幼芽及び幼根の伸長に対する低線量照射の影響をまとめると、幼芽への影響は幼根への影響に比べれば少ない。幼根への影響は、品種の差が余りにも大きく、しかも品種の異なる試料の前処理条件にも差があり、統一的結論は出し難い。
|
 |
米の根端の細胞は小麦のそれに比べ小さく、ハイゲンハイン氏ハマトキシリン染色によって細胞分裂中期および後期は良く染色されるが、前期および終期の完全な染色は困難であった。非照射群にあっては、細胞分裂後期におけるブリッジの存在は稀であるが、照射群では数本のブリッジが観察された。しかしながら、10~30kradの照射範囲ではdose responseは見られず、また小核の存在も認められなかった。
「日本晴」を試料とした根端140μm位における細胞分裂異常の出現率を表6-2に示した。
短期間貯蔵穀粒(2ヶ年以内)にあっては、発芽期の根端における細胞分裂後期細胞に出現するブリッジの発現頻度を指標として、照射処理の有無を検知し得ると思われる。しかし、発芽し得ない試料、たとえば長期間(2年以上)貯蔵したもの、人工乾燥などによって細胞が
| 照 射 量 (krad) |
試料粒 |
総細胞数 |
中期細胞 |
後期細胞 |
ブリッジ を有する 後期細胞 |
後期細胞 中の割合 (%) |
| 0 10 20 30 |
10 10 10 10 |
1,790 2,085 1,888 1,749 |
16 22 21 19 |
17 14 18 13 |
1 11 16 11 |
5.9 78.6 88.9 84.6 |