Fig. 1 Alkaline comet assay of cherry seed embryos.
 |
幚尡曽朄
丂1. 帋椏媦傃徠幩
丂2. DNA 僐儊僢僩傾僢僙僀
丂3. 僴乕僼僄儞僽儕僆僥僗僩
幚尡寢壥媦傃峫嶡
丂1. 僐儊僢僩傾僢僙僀偵傛傞僒僋儔儞儃泱偺 DNA 懝彎偺娤嶡
丂2. 僈儞儅慄徠幩偺敪夎偵媦傏偡塭嬁
丂専塽張棟傗搚忞徚撆偵梡偄傜傟偰偄傞廘壔儊僠儖偼丄僆僝儞憌攋夡暔幙偱偁傞偲偟偰偦偺巊梡偑 2005 擭傪婜尷偵慡攑偝傟傞偙偲偑寛掕偟偰偄傞丅曻幩慄徠幩偼丄専塽張棟偵偍偗傞廘壔儊僠儖鄮忲偺戙懼張棟偲偟偰桳朷帇偝傟偰偍傝丄庬乆偺擾嶻暔偵偍偗傞専塽懳徾奞拵偵懳偡傞桳岠側曻幩慄偺慄検偺専摙偑峴傢傟偰偄傞1)丅
丂専塽張棟偵曻幩慄徠幩傪摫擖偡傞偨傔偵偼丄偦偺揔惓側棳捠傪娗棟偡傞偨傔偵丄壥幚傗擾嶻暔傪懳徾偵偟偨掅慄検偺徠幩偺桳柍傪専弌偡傞専抦媄弍偺妋棫偑朷傑傟傞丅偙傟傑偱偵丄壨懞傜偼丄壥幚庬巕偺敪夎擻椡傪恦懍娙曋偵應掕偡傞丄僴乕僼僄儞僽儕僆僥僗僩朄傪奐敪偟偰丄姽媖椶2) 傗儕儞僑媦傃僒僋儔儞儃3) 偺専抦偵墳梡偟丄傑偨丄偙偺曽朄偺桳岠惈傪暋悢帋尡婡娭偵偍偗傞僽儔僀儞僪僥僗僩傪巊偭偰専徹偟偰偄傞4)丅
丂DNA 僐僢儊僩傾僢僙僀偼丄曻幩慄側偳偵傛偭偰婲偙傞 DNA 偺懝彎傪専弌偡傞媄弍偱丄徠幩怘昳偺娙曋側僗僋儕乕僯儞僌朄偲偟偰摦暔惈怘昳傗怉暔庬巕偵墳梡偝傟偰偄傞5) 乣 9)丅偙偺曽朄偼丄曻幩慄偵傛傞 DNA 懝彎 乮嵔愗抐乯 傪崅姶搙偵専弌偱偒傞偨傔丄尨棟揑偵偼丄DNA 傪娷傓摦怉暔怘昳偵懳偟偰峀斖偵墳梡偱偒傞壜擻惈傪桳偟偰偄傞丅
丂尰嵼丄擔杮崙撪偵桝擖偝傟偰偄傞壥幚偺拞偱丄専塽偵偍偗傞徚撆朄偵廘壔儊僠儖鄮忲偑 100% 嵦梡偝傟偰偄傞傕偺偺堦偮偲偟偰丄僇僫僟丄傾儊儕僇丄僯儏乕僕乕儔儞僪嶻偺僒僋儔儞儃偑偁傞丅僒僋儔儞儃偺専塽懳徾奞拵偺僐僪儕儞僈偺徚撆偵桳岠側僈儞儅慄偺慄検偼 140Gy 偲曬崘偝傟偰偍傝10)丄僒僋儔儞儃庬巕偺僴乕僼僄儞僽儕僆僥僗僩偺専弌尷奅偼丄150Gy 偲曬崘偝傟偰偄傞偙偲偐傜丄偙偺曽朄偼丄僒僋儔儞儃偵懳偡傞幚梡揑側徠幩偺専弌偵懳墳偱偒傞傕偺偲敾抐偝傟傞丅偟偐偟丄敾掕偵梫偡傞帪娫偑 4 擔偲挿偄偨傔丄傛傝抁婜娫偱敾掕偺偱偒傞曽朄偺奐敪偑朷傑傟傞丅
丂杮尋媶偱偼丄専塽偵梫偡傞慄検傪憐掕偟偰丄僒僋儔儞儃壥幚傪 100Gy-200Gy 徠幩偟丄庬巕泱偺嵶朎 DNA 偺懝彎傪僐儊僢僩傾僢僙僀偵傛傝應掕偟偨丅摨帪偵僴乕僼僄儞僽儕僆僥僗僩傪幚巤偟偰椉幰偺斾妑傪峴偄丄専抦媄弍偲偟偰偺壜擻惈傪媍榑偡傞偲摨帪偵丄僈儞儅慄徠幩偵傛傞敪夎擻椡偺寚懝偲 DNA 嵔愗抐媦傃廋暅擻偲偺娭學偵偮偄偰峫嶡偟偨丅
丂僒僋儔儞儃 乮傾儊儕僇嶻乯 偼巗斕偺傕偺傪梡偄偨丅壥幚偺徠幩偼丄僈儞儅僙儖 220 乮慄検棪 1.5kGy/h) 傪梡偄丄100丄150 偍傛傃 200Gy 偺慄検偱忢壏偵偰峴偭偨丅 徠幩屻偺壥幚偼 5亷 偺椻憼屔偵曐懚偟偨丅
丂僒僋儔儞儃偺庬巕偺奜庬旂傪攋嵱丄撪庬旂傪庢傝彍偄偨屻偵泱傪庢傝弌偟偨丅偙傟偵塼懱拏慺傪壛偊偰媫椻偟丄搥寢忬懺偺泱傪擕敨偱暡嵱偟偨丅泱 1 屄偵偮偒 2ml 偺 25mM EDTA 傪娷傓 PBS 娚徴塼拞傪壛偊丄昘拞偱 5 暘潣漚偟丄50兪m 偺僫僀儘儞儊僢僔儏偱傠夁偟丄5 暘娫惷抲偟偨忋惔偐傜妀抐曅傪廤傔偨丅偙偺帋椏偵偮偄偰丄Sigh 傜偺11) 曽朄偵婎偯偄偰丄傾儖僇儕忦審偱僐儊僢僩傾僢僙僀傪峴偭偨丅偡側傢偪丄妀抐曅傪 10 攞検偺 0.5% 偺掅梈揰傾僈儘乕僗偲崿崌偟偨屻丄僗儔僀僪僌儔僗忋偵屌掕偟偨丅偙傟傪丄2.5M NaCl丄1% 僒儖僐僔儖僫僩儕僂儉丄100mM EDTA 10mM Tris (pH=10.0) 1% Triton X-100丄 10% DMSO 偐傜側傞 lysis 梟塼偵偮偗 1 帪娫 5亷 偱曻抲偟丄僞儞僷僋幙傪梟夝偟偨丅僒僽儅儕儞揹婥塲摦憛偵 300mM NaOH 1mM EDTA (pH亜13) 傪枮偨偟丄嵶朎傪屌掕偟偨僗儔僀僪僌儔僗傪暲傋丄20 暘娫曻抲偟偰丄unwinding 傪峴偭偨偺偪丄0.75V/cm 偺忦審偱 15 暘娫塲摦偟偨丅
丂僐儊僢僩憸偼 20兪g/ml 偺儓僂壔僾儘僺儖偱 3 暘娫愼怓偟偨偺偪丄2 暘娫悈愻偟丄寀岝尠旝嬀 乮OLYMPUS BX50乯 偱娤嶡偟偨丅柍嶌堊偵帇栰傪慖傫偱丄1 僒儞僾儖偁偨傝 20 屄埲忋偺僐儊僢僩憸傪丄CCD 僇儊儔 乮MITSUBISHI CIT605乯 傪梡偄偰嶣塭偟丄僐儞僺儏乕僞夋憸偲偟偰曐懚偟偨丅僐儊僢僩夋憸偺夝愅偼夋憸夝愅僜僼僩 乮儐乕儚乕僋僗惢丄http://www.youworks.co.jp/乯 傪梡偄偰峴偄丄Tail Moment 傪寁嶼偟偨丅
丂壨懞傜偺曽朄3) 偵廬偭偰峴偭偨丅僈儞儅慄徠幩屻 13 擔娫椻憼曐懚偟偨壥幚偐傜丄僐儊僢僩傾僢僙僀偺帪偲摨條偵泱傪摼偨丅偙傟偺泱偺巕梩傪 1 枃彍嫀偟偰僴乕僼僄儞僽儕僆傪挷惢偟偨丅奺張棟嬫 10 屄偺僴乕僼僄儞僽儕僆傪僔儍乕儗拞偺 10兪M 儀儞僕儖傾僨僯儞梟塼傪怹偟偨扙帀柸忋偵抲偒丄傆偨傪偟偰埫強偵偍偄偰 25亷 偱攟梴偟丄敪夎媦傃敪崻偺條巕傪娤應偟偨丅
丂徠幩捈屻偵僒僋儔儞儃庬巕偺泱偐傜妀抐曅傪挷惢偟丄傾儖僇儕忦審偱僐儊僢僩傾僢僙僀傪峴偆偲丄徠幩僒儞僾儖偱偼丄DNA 嵔愗抐偵桼棃偡傞挿偔旜傪堷偄偨宍偺僐儊僢僩憸偑娤應偝傟偨丅傑偨丄旕徠幩偺僒儞僾儖偱偼丄傎傏媴宍偺塲摦憸偑娤應偝傟丄DNA 嵔偺愗抐偼傎偲傫偳娤應偝傟側偐偭偨丅傑偨丄壥幚偺挋憼偵傛傞塭嬁傪専摙偟偨偲偙傠丄徠幩 36 帪娫屻偵偼丄徠幩僒儞僾儖偺僐儊僢僩憸偺 Tail 偑抁偔側傞偺偑娤嶡偝傟丄72 帪娫屻偵偼丄200Gy 偵偍偄偰傕懝彎偵傛傞 Tail 偑傎偲傫偳専弌偝傟偢丄旕徠幩偲摨條偺媴宍偺塲摦憸偑娤應偝傟偨 (Fig. 1)丅
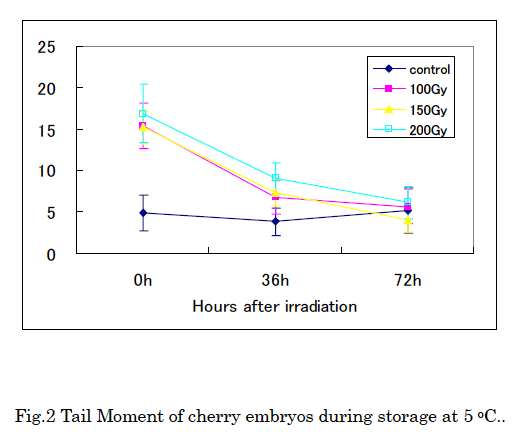
丂夋憸夝愅憰抲傪梡偄偰丄懝彎偺巜昗偱偁傞 Tail Moment 乮DNA 偺堏摦搙偲懝彎晹偺寀岝嫮搙偺僐儊僢僩慡懱偺寀岝偵懳偡傞斾棪偺愊嶼乯 傪寁嶼偟丄挋憼拞偺曄壔傪捛愓偟偨寢壥傪 Fig. 2 偵帵偡丅徠幩捈屻偼丄偳偺慄検偵偍偄偰傕丄旕徠幩偺僐儞僩儘乕儖偵斾傋偰 Tail Moment 偺桳堄側憹壛偑尒傜傟偨偑丄72 帪娫屻偵偼丄200Gy 徠幩偺僒儞僾儖偱傕僐儞僩儘乕儖偲偺嵎偼柍偔側偭偨丅
丂傾儖僇儕忦審偺僐儊僢僩傾僢僙僀偱偼丄DNA 偵惗偠偨 1 杮嵔愗抐偲 2 杮嵔愗抐偺椉曽傪専弌偟偰偄傞丅徠幩捈屻偵偼丄偙傟傜偺 DNA 嵔愗抐偵桼棃偡傞懝彎偺戝偒偄僐儊僢僩憸偑娤嶡偝傟偨偑丄徠幩屻偺挋憼拞偵嵔愗抐偺廋暅偑峴傢傟丄36 帪娫屻偵偼 Tail 偺挿偝偑抁偔側傝 72 帪娫屻偵偼丄嵔愗抐偼廋暅偝傟偨偲峫偊傜傟傞丅姰弉偟偨姡憞庬巕偵偍偄偰偼丄DNA 懝彎偺廋暅偼丄媧悈屻敪夎帪偵婲偙傞偲曬崘偝傟偰偍傝12), 13)丄Koppenn 傜偼丄僟僀僗丄傂傑傢傝庬巕側偳悢庬偺姡憞忬懺偺怉暔庬巕偵偮偄偰丄拞惈偺僐儊僢僩傾僢僙僀傪梡偄偰丄徠幩偺桳柍傪 0.1kGy 傑偱専弌偱偒傞偲曬崘偟偰偄傞6)丅傑偨丄Barros 傜傕彫敒偵偮偄偰 6 儢寧娫偺挋憼婜娫傪宱偰傕 0.1kGy 偺徠幩偺棜楌傪僐儊僢僩傾僢僙僀偵傛偭偰専弌偱偒傞偲曬崘偟偰偄傞8)丅偟偐偟丄崱夞変乆偑峴偭偨丄僒僋儔儞儃壥幚拞偺姡憞偟偰偄側偄庬巕泱偱偼丄廋暅懍搙偑懍偔丄200Gy 偺徠幩偱傕棳捠婜娫偵偍偗傞徠幩偺桳柍偺専弌偵僐儊僢僩傾僢僙僀傪梡偄傞偙偲偼丄擄偟偄偲敾抐偝傟偨丅
|
 |
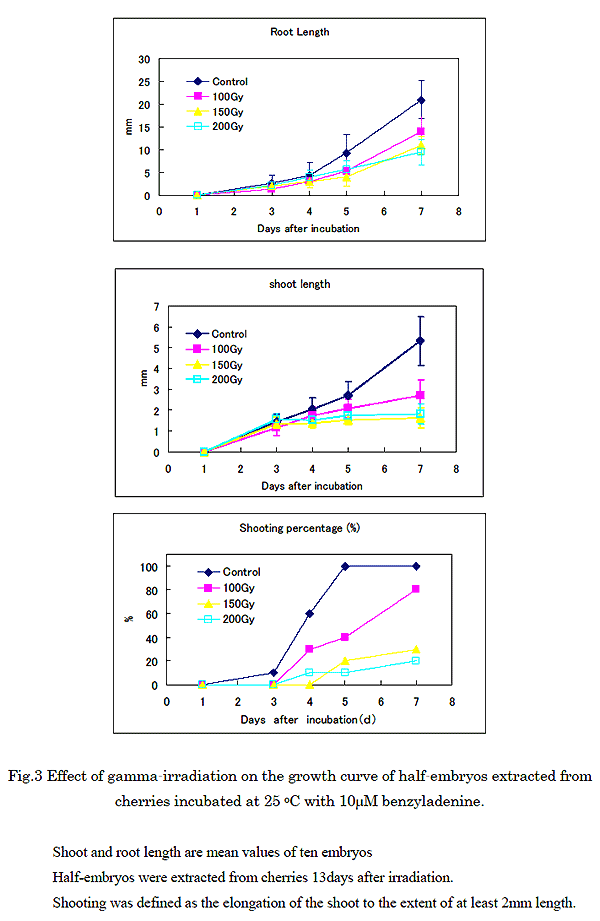
丂Fig. 3 偵徠幩偟偨僒僋儔儞儃壥幚傪 13 擔娫丄椻憼偟偨屻僴乕僼僄儞僽儕僆僥僗僩傪挷惢偟丄25亷 偱攟梴偟偨帪偺夎偲崻偺挿偝傪應掕偟偨寢壥傪帵偡丅傑偨丄Fig. 4 偵攟梴 5 擔屻偺僴乕僼僄儞僽儕僆偺幨恀傪帵偡丅攟梴 3 擔屻偵偼丄偄偢傟偺張棟嬫偵偍偄偰傕丄傢偢偐側敪崻丄敪夎偑擣傔傜傟丄旕徠幩偺僐儞僩儘乕儖偱偼丄攟梴擔悢偺宱夁偲偲傕偵丄夎丄崻偲傕偵怢挿偑擣傔傜傟偨丅堦曽丄僈儞儅慄張棟傪峴偭偨壥幚偐傜拪弌偟偨庬巕偱偼丄崻媦傃夎偺怢挿偑慄検偵墳偠偰抶傟傞偺偑擣傔傜傟偨丅摿偵丄徠幩嬫偵偍偗傞敪夎偺抶傟偼尠挊偱丄150Gy 埲忋偺徠幩偱偼丄攟梴 7 擔屻偱傕丄夎偺挿偝偺暯嬒抣偑 2mm 傪墇偊傞偙偲偑柍偐偭偨丅壨懞傜偼丄僒僋儔儞儃偵偍偗傞僴乕僼僄儞僽儕僆帋尡偺曽朄偲偟偰丄夎偺挿偝 1mm 埲忋傪敪夎偲掕媊偟偰丄10 屄懱埲忋偺僴乕僼僄儞僽儕僆偺敪夎棪偑攟梴 4 擔屻偵 50% 偵払偟側偄傕偺傪徠幩偲敾掕偡傞偲曬崘偟偰偄傞4)丅偟偐偟丄杮幚尡偱偼丄徠幩嬫偵偍偄偰傕丄攟梴 3 擔屻偐傜傢偢偐偱偼偁傞偑夎偺怢挿偑擣傔傜傟偨偨傔丄敪夎偺掕媊傪夎偺挿偝偑 2mm 埲忋偵払偟偨傕偺偲偟丄敪夎棪偺宱擔揑曄壔傪僌儔僼偵帵偟偨丅旕徠幩偺僐儞僩儘乕儖偱偼丄攟梴 4 擔屻偵敪夎棪偑 60%丄5 擔屻偵偼 100% 偵払偟偨丅堦曽丄100Gy 徠幩偱偼丄攟梴 5 擔屻偱傕敪夎棪偑 50% 傪墇偊偢丄7 擔屻偵側偭偰丄80% 偵摓払偟偨丅150Gy 埲忋偺徠幩偱偼丄攟梴 7 擔屻偵側偭偰傕丄敪夎棪偼 20% 埲壓偲掅偄傑傑偱偁偭偨丅埲忋偐傜徠幩屻偺挋憼偵娭傢傜偢丄攟梴 4 擔屻偺敪夎棪丄50% 傪婎弨偲偟丄100Gy 傪尷奅偲偟偰徠幩偺桳柍偺敾暿偑壜擻偱偁偭偨丅
丂偙傟傜偺寢壥偐傜丄僐儊僢僩傾僢僙僀偱専弌壜擻側 DNA 嵔偺愗抐偑挋憼婜娫拞偵廋暅偝傟偰傕丄敪夎擻椡偼夞暅偣偢丄専塽張棟傪憐掕偟偨掅慄検偺僈儞儅慄徠幩偺桳柍偺専弌偑丄敪夎帋尡 乮僴乕僼僄儞僽儕僆乯 帋尡偱壜擻偁傞偲寢榑偝傟偨丅
 |
 |
1) G. J. Hallman : in Food Irradiation: Principles and Applications, Ed by R. A. Molis (John Wiley & Sons Inc.) pp.113-130.
2) Y. Kawamura, S. Uchiyama, Y. Saito : J Food Sci., 54 (6), 1501-1504(1989).
3) Y. Kawamura, A. Miura, T. Sugita, T. Yamada, Y. Saito : Radiat. Phys. Chem, 46 (3), 371亅375 (1995).
4) Y. Kawamura, T. Sugita, T. Yamada, Y. Saito : Radiat. Phys. Chem., 48 (5), 665-668 (1996).
5) H. Cerda, H. Delincee, H. Haine, H. Rupp : Mutation Research : 375, 167-181 (1997).
6) G. Koppen, H. Cerda : Lebensm.-Wiss.u.-Technol., 30, 452-457 (1997).
7) A. A. Khan, H. M. Khan, H. Delincee : J Food Sci., 67 (2), 493-496 (2002).
8) A. C. Barros, M. T. L. Freund, A. L. C. H. Villavicencio, H. Delincee, V. Arthur : Radiat. Phys. Chem., 63, 423-426 (2002).
9) European Committee for Standardization EN 13784 (2001).
10) IAEA : IDIDAS僨乕僞儀乕僗 http://www-ididas.iaea.org/IDIDAS/start.htm
11) Singh, N. P., McCoy, M. T., Tice, R. R., Schneider, E. L., Experimental Cell Research 175, 184 (1988).
12) R. H. Elder et. al : Mutat Res. 181, 61-71 (1987).
13) R. H. Elder and D. J. Osborne : Seed Sci. Res. 3, 43-53 (1993).
乮2002 擭 8 寧 2 擔庴棟乯
|
|